

Opsonization modulates Rac-1 activation during cell entry by Leishmania amazonensis
- PMID: 12117970
- PMCID: PMC128177
- DOI: 10.1128/IAI.70.8.4571-4580.2002
Opsonization modulates Rac-1 activation during cell entry by Leishmania amazonensis
Abstract
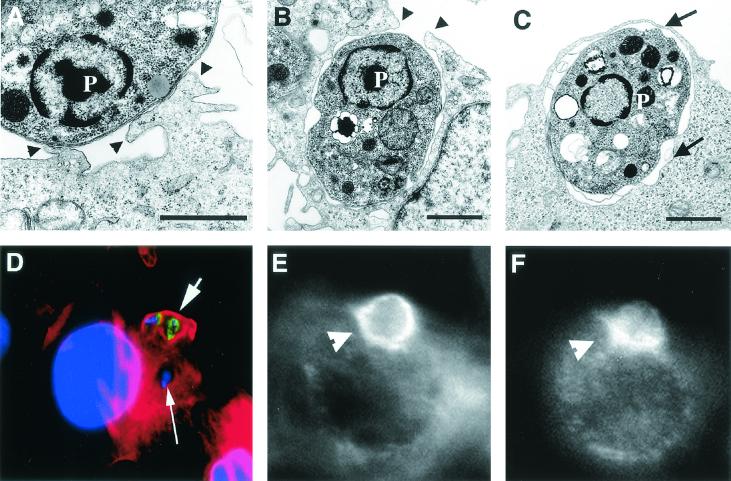
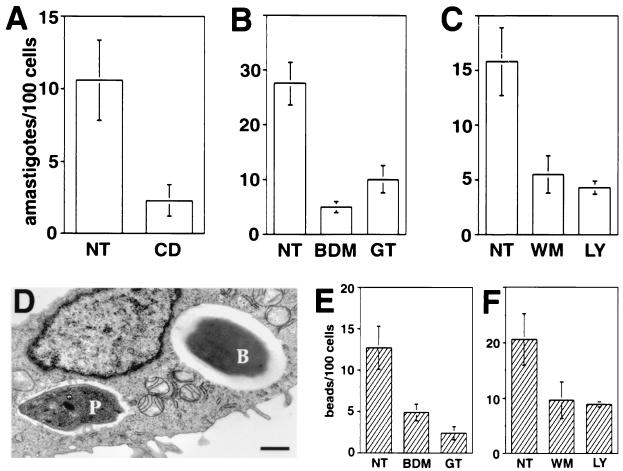
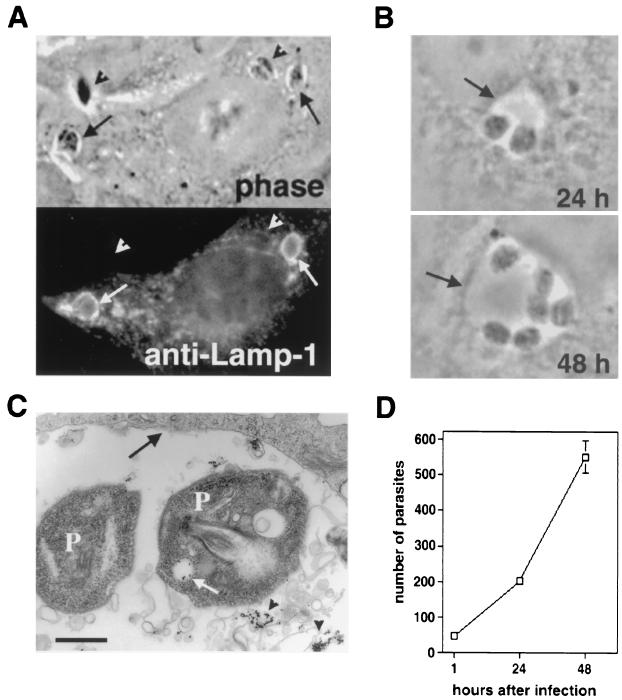
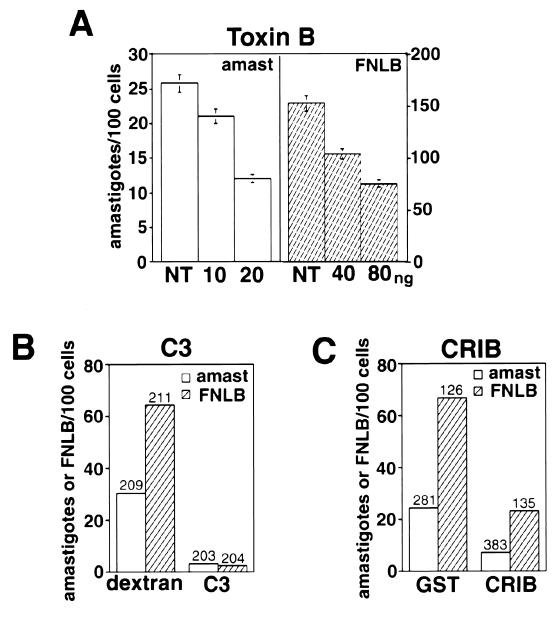
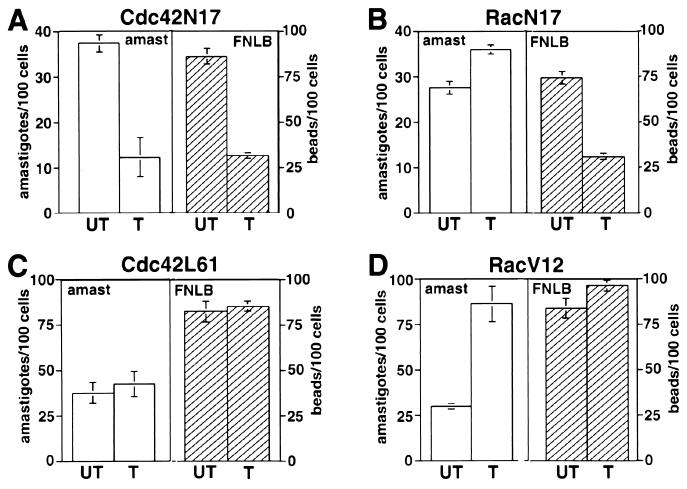
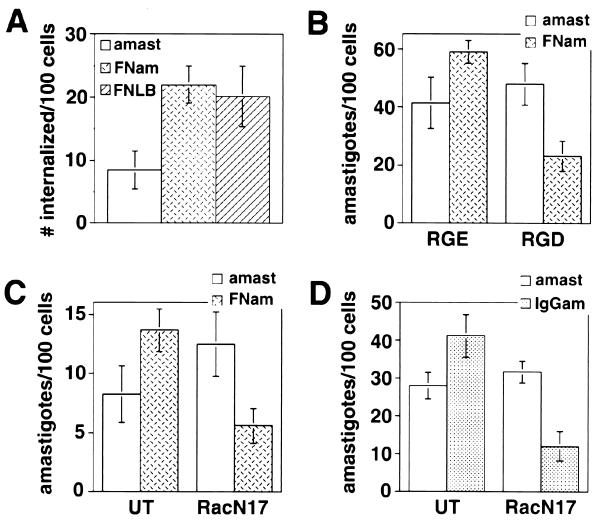
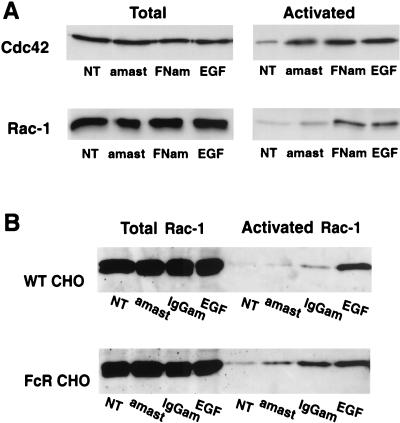
Lesions caused by Leishmania amazonensis normally heal, but relapses occur due to parasite persistence in host tissues. It has been proposed that infection of fibroblasts plays an important role in this process by providing the parasites with a safe haven in which to replicate. However, most previous studies have focused on the entry of Leishmania into macrophages, a process mediated by serum opsonins. To gain insight into a possible role of nonopsonic entry in the intracellular persistence of amastigotes, we examined the invasion of Chinese hamster ovary (CHO) cells. Amastigotes entered CHO cells by a cytochalasin D, genistein, wortmannin, and 2,3-butanedione monoxime-sensitive pathway and replicated within phagolysosomes. However, unlike most phagocytic processes described to date, amastigote internalization in CHO cells involved activation of the GTPases Rho and Cdc42 but not Rac-1. When uptake was mediated by fibronectin or when amastigotes were opsonized with immunoglobulin G and internalized by Fc receptor-expressing CHO cells, Rac-1 activation was restored and found to be required for parasite internalization. Given the essential role of Rac in assembly of the respiratory burst oxidase, invasion through this nonopsonic, Rac-1-independent pathway may play a central role in the intracellular survival of Leishmania in immune hosts.
Figures
Similar articles
-
Leishmania amazonensis: the phagocytosis of amastigotes by macrophages.Exp Parasitol. 1998 Mar;88(3):161-71. doi: 10.1006/expr.1998.4232. Exp Parasitol. 1998. PMID: 9562419
-
Immune serum from both susceptible and resistant strains of mice increases phagocytosis of Leishmania mexicana amazonensis by macrophages.Acta Trop. 1987 Sep;44(3):339-42. Acta Trop. 1987. PMID: 2892370
-
Phagocytosis of Leishmania donovani amastigotes is Rac1 dependent and occurs in the absence of NADPH oxidase activation.Eur J Immunol. 2006 Oct;36(10):2735-44. doi: 10.1002/eji.200636089. Eur J Immunol. 2006. PMID: 16955522
-
Leishmania amazonensis: multiple receptor-ligand interactions are involved in amastigote infection of human dendritic cells.Exp Parasitol. 2007 Jul;116(3):306-10. doi: 10.1016/j.exppara.2007.01.003. Epub 2007 Jan 19. Exp Parasitol. 2007. PMID: 17320869
-
Modular components of phagocytosis.J Leukoc Biol. 1999 Nov;66(5):712-7. doi: 10.1002/jlb.66.5.712. J Leukoc Biol. 1999. PMID: 10577498 Review.
Cited by
-
The Abl and Arg kinases mediate distinct modes of phagocytosis and are required for maximal Leishmania infection.Mol Cell Biol. 2012 Aug;32(15):3176-86. doi: 10.1128/MCB.00086-12. Epub 2012 Jun 4. Mol Cell Biol. 2012. PMID: 22665498 Free PMC article.
-
The Leishmania surface protease GP63 cleaves multiple intracellular proteins and actively participates in p38 mitogen-activated protein kinase inactivation.J Biol Chem. 2009 Mar 13;284(11):6893-908. doi: 10.1074/jbc.M805861200. Epub 2008 Dec 8. J Biol Chem. 2009. PMID: 19064994 Free PMC article.
-
Role of Host Small GTPases in Apicomplexan Parasite Infection.Microorganisms. 2022 Jul 7;10(7):1370. doi: 10.3390/microorganisms10071370. Microorganisms. 2022. PMID: 35889089 Free PMC article. Review.
-
Using machine learning to dissect host kinases required for Leishmania internalization and development.bioRxiv [Preprint]. 2024 Aug 4:2024.05.16.593986. doi: 10.1101/2024.05.16.593986. bioRxiv. 2024. Update in: Mol Biochem Parasitol. 2024 Dec;260:111651. doi: 10.1016/j.molbiopara.2024.111651. PMID: 38798624 Free PMC article. Updated. Preprint.
-
Oxidant generation by single infected monocytes after short-term fluorescence labeling of a protozoan parasite.Infect Immun. 2007 Feb;75(2):1017-24. doi: 10.1128/IAI.00914-06. Epub 2006 Nov 21. Infect Immun. 2007. PMID: 17118986 Free PMC article.
References
-
- Alexander, J., A. R. Satoskar, and D. G. Russell. 1999. Leishmania species: models of intracellular parasitism. J. Cell Sci. 112:2993-3002. - PubMed
-
- Antoine, J. C., E. Prina, T. Lang, and N. Courret. 1998. The biogenesis and properties of the parasitophorous vacuoles that harbour Leishmania in murine macrophages. Trends Microbiol. 6:392-401. - PubMed
-
- Arlein, W. J., J. D. Shearer, and M. D. Caldwell. 1998. Continuity between wound macrophage and fibroblast phenotype: analysis of wound fibroblast phagocytosis. Am. J. Physiol. 275:R1041-R1048. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
