

Mobilization of protein kinase C in macrophages induced by Listeria monocytogenes affects its internalization and escape from the phagosome
- PMID: 12117979
- PMCID: PMC128209
- DOI: 10.1128/IAI.70.8.4650-4660.2002
Mobilization of protein kinase C in macrophages induced by Listeria monocytogenes affects its internalization and escape from the phagosome
Abstract
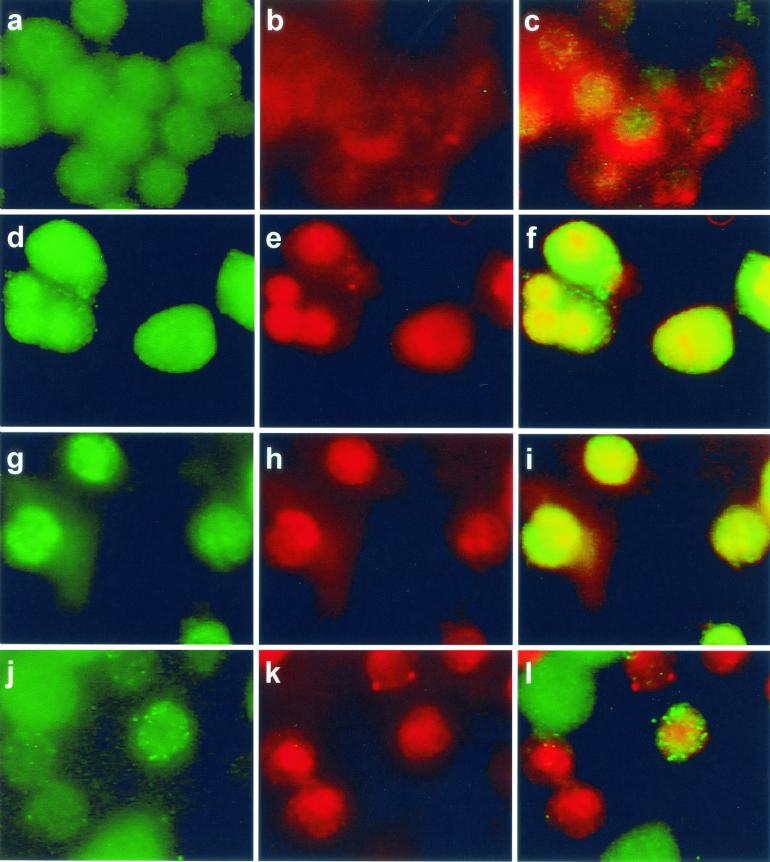
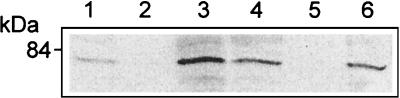
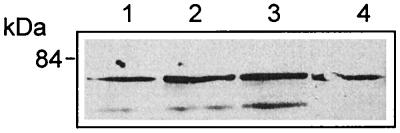
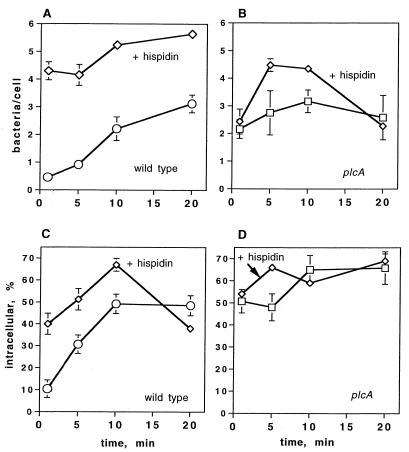
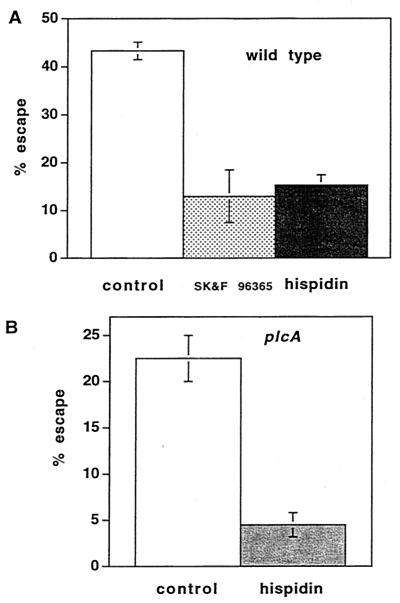
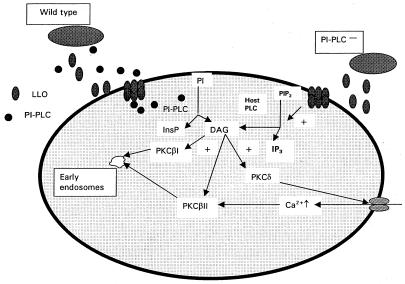
Listeriolysin O (LLO) and a phosphatidylinositol-specific phospholipase C (PI-PLC) are known virulence factors of Listeria monocytogenes in both tissue cultures and the murine model of infection. LLO is a member of a family of pore-forming cholesterol-dependent cytotoxins and is known to play an essential role in escape from the primary phagocytic vacuole of macrophages. PI-PLC plays an accessory role, in that PI-PLC mutants are partially defective in escape. We have shown that both of these molecules are essential for initiating rapid increases in the calcium level in the J774 murine macrophage cell line (S. J. Wadsworth and H. Goldfine, Infect. Immun. 67:1770-1778, 1999). Here we show that both LLO and PI-PLC are required for translocation of protein kinase C delta (PKC delta) to the periphery of J774 cells and for translocation of PKC beta II to early endosomes beginning within the first minute after addition of bacteria to the culture medium. Treatment with the calcium channel blocker SK&F 96365 inhibited translocation of PKC beta II but not PKC delta. Our findings lead us to propose a host signaling pathway requiring LLO and the formation of diacylglycerol by PI-PLC in which calcium-independent PKC delta is responsible for the initial calcium signal and the subsequent PKC beta II translocation. LLO-dependent translocation of PKC beta I to early endosomes also occurs between 1 and 4 min after infection, but this occurs in the absence of PI-PLC. All of these signals were observed in cells that had not internalized bacteria. Blocking PKC beta translocation with hispidin resulted in more rapid uptake of wild-type bacteria and greatly reduced escape from the primary phagocytic vacuoles of J774 cells.
Figures
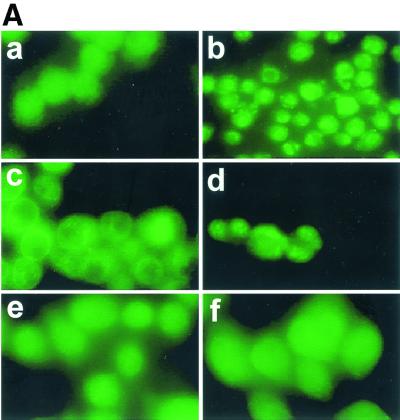
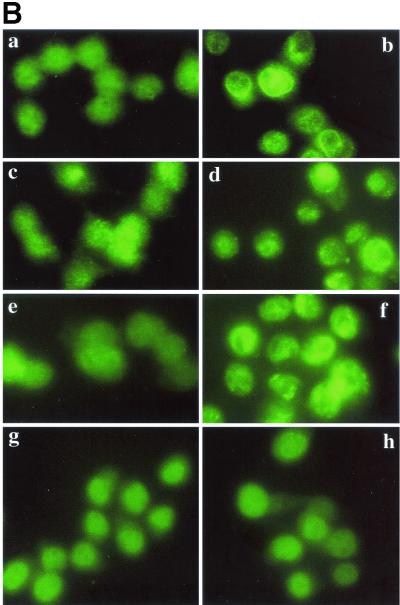
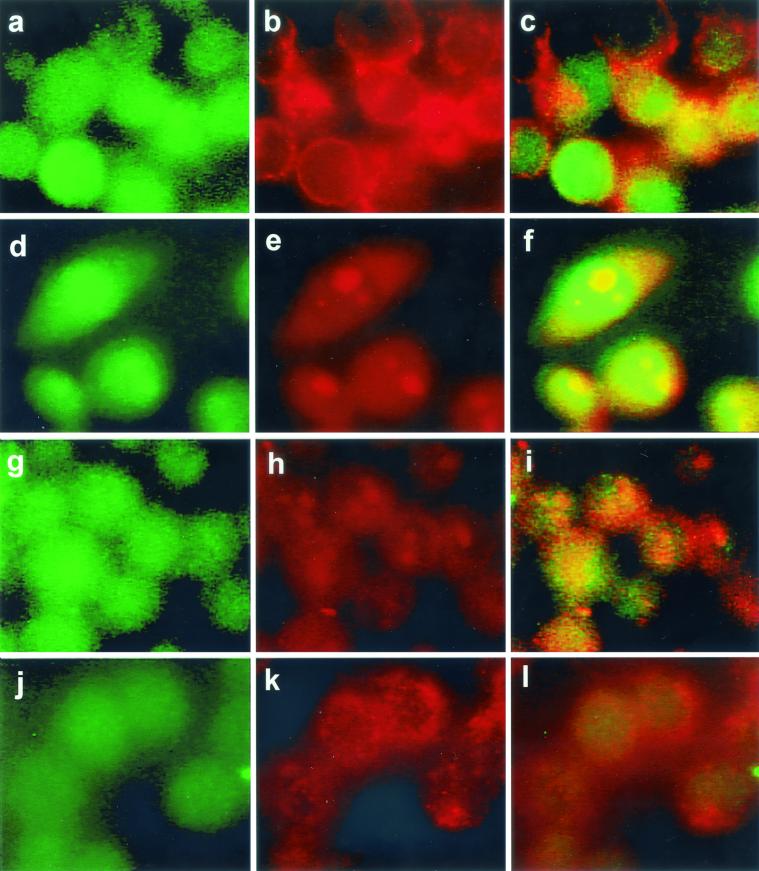
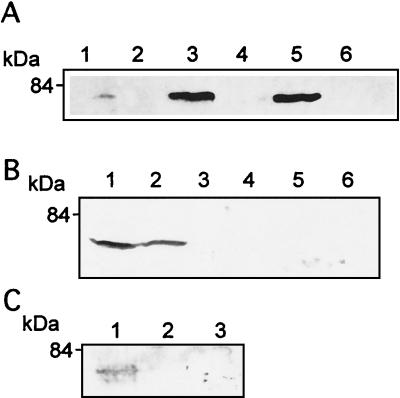
References
-
- Alvarez-Dominguez, C., A. M. Barbieri, W. Berón, A. Wandinger-Ness, and P. D. Stahl. 1996. Phagocytosed live Listeria monocytogenes influences rab5-regulated in vitro phagosome-endosome fusion. J. Biol. Chem. 271:13834-13843. - PubMed
-
- Bubert, A., Z. Sokolovic, S. K. Chun, L. Papatheodorou, A. Simm, and W. Goebel. 1999. Differential expression of Listeria monocytogenes virulence genes in mammalian host cells. Mol. Gen. Genet. 261:323-336. - PubMed
-
- Bucci, C., R. G. Parton, I. H. Mather, H. Stunnenberg, K. Simons, B. Hoflack, and M. Zerial. 1992. The small GTPase rab5 functions as a regulatory factor in the early endocytic pathway. Cell 70:715-728. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
