

CD4(+)CD25(+) regulatory T cells can mediate suppressor function in the absence of transforming growth factor beta1 production and responsiveness
- PMID: 12119348
- PMCID: PMC2193919
- DOI: 10.1084/jem.20020590
CD4(+)CD25(+) regulatory T cells can mediate suppressor function in the absence of transforming growth factor beta1 production and responsiveness
Abstract
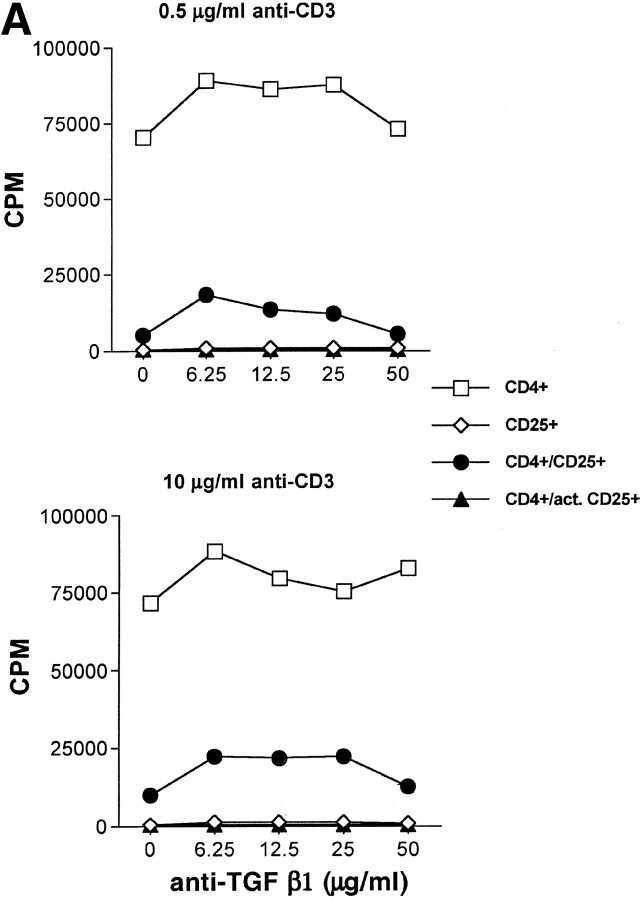
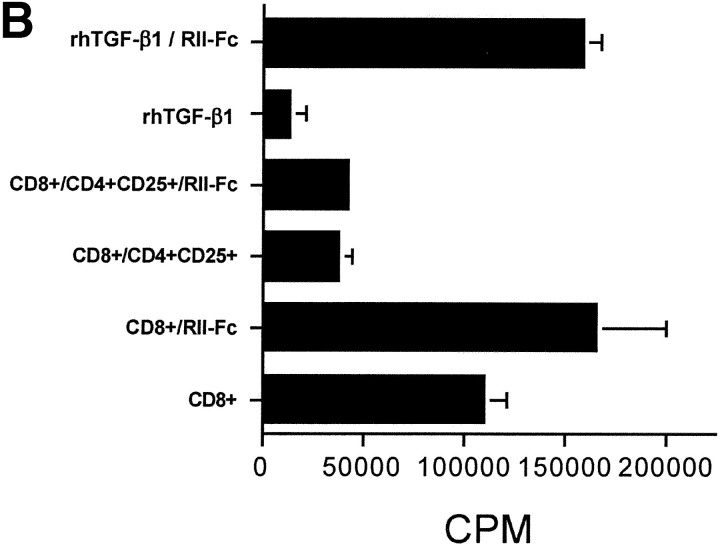
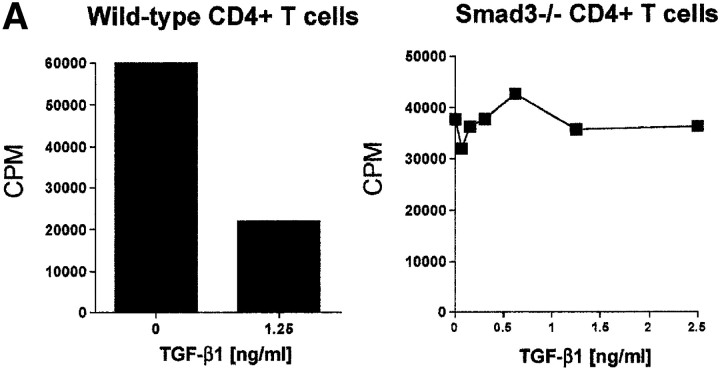
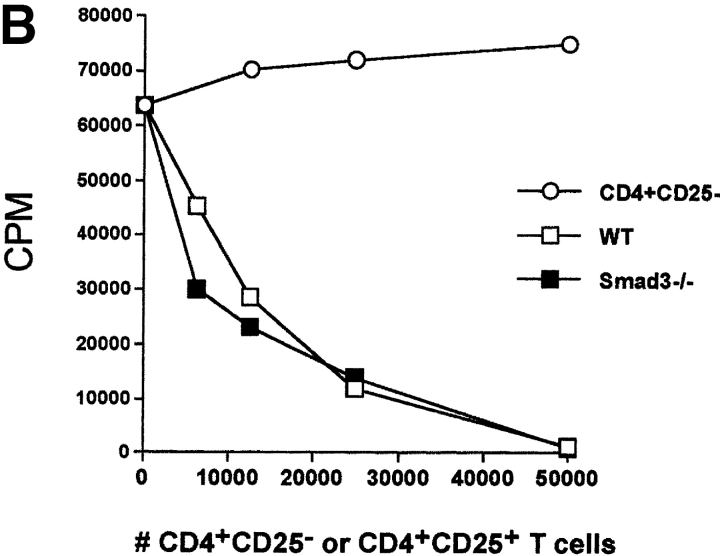
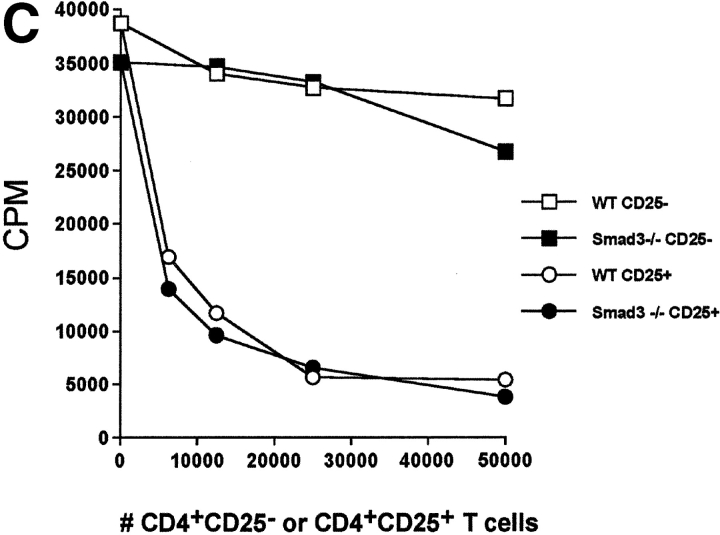
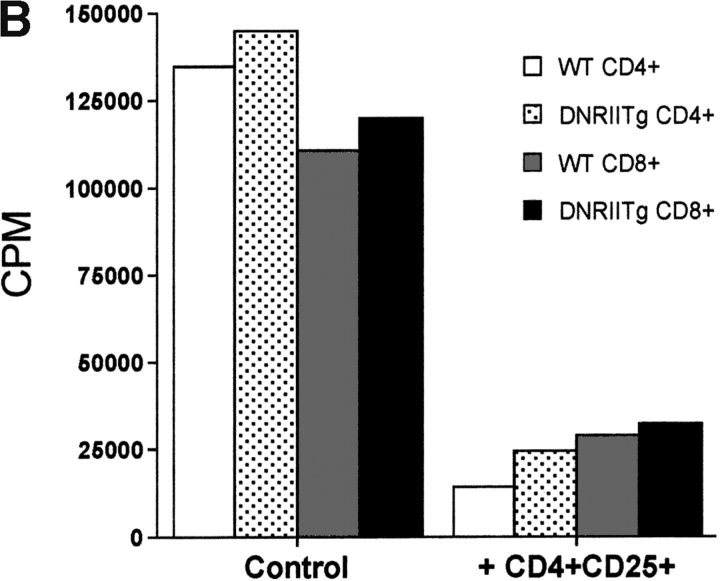
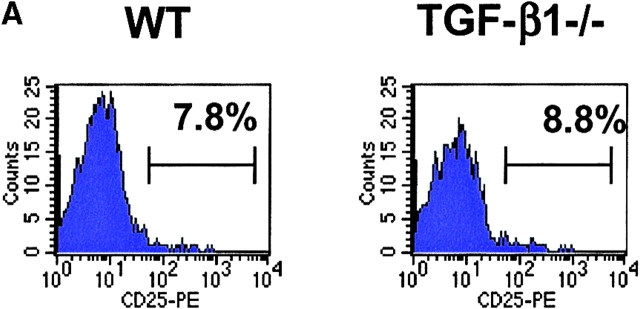
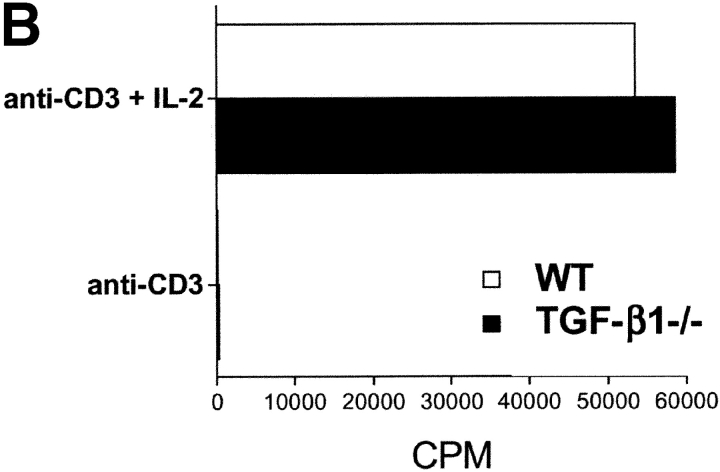
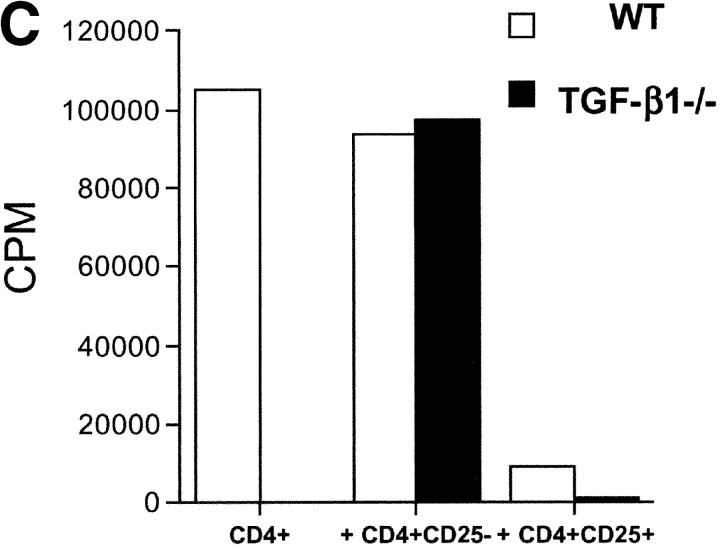
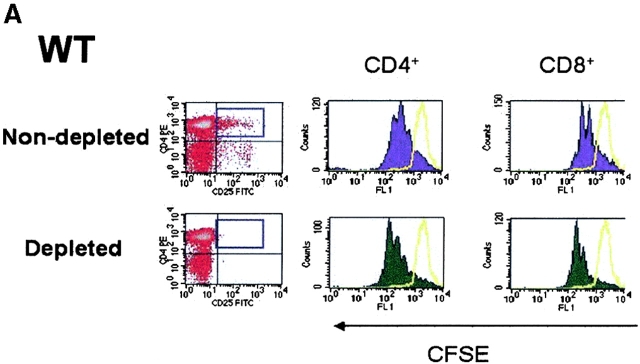
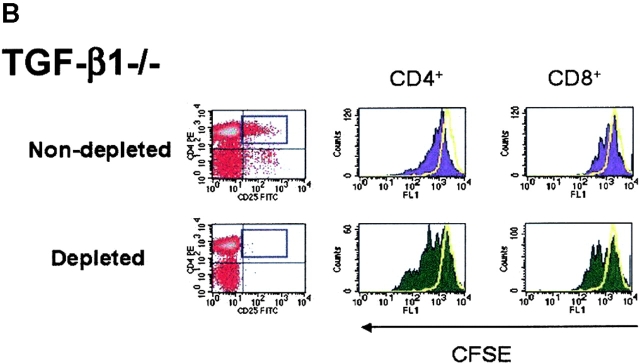
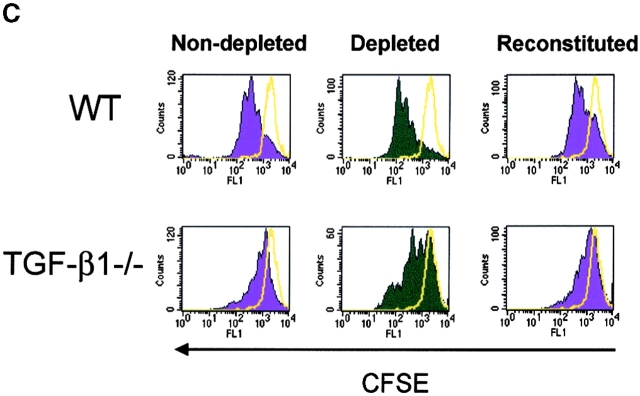
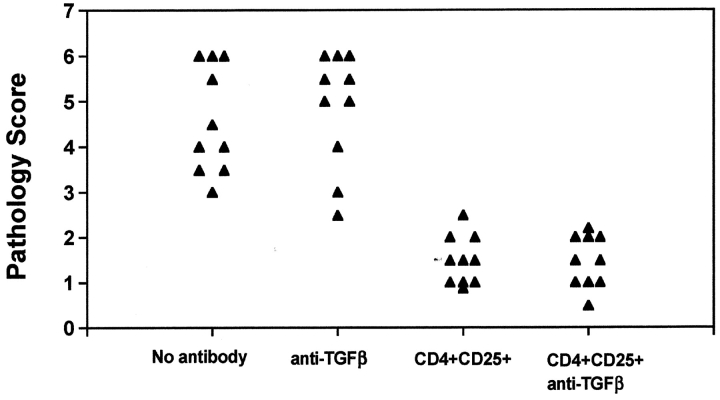
CD4(+)CD25(+) regulatory T cells inhibit organ-specific autoimmune diseases induced by CD4(+)CD25(-) T cells and are potent suppressors of T cell activation in vitro. Their mechanism of suppression remains unknown, but most in vitro studies suggest that it is cell contact-dependent and cytokine independent. The role of TGF-beta1 in CD4(+)CD25(+) suppressor function remains unclear. While most studies have failed to reverse suppression with anti-transforming growth factor (TGF)-beta1 in vitro, one recent study has reported that CD4(+)CD25(+) T cells express cell surface TGF-beta1 and that suppression can be completely abrogated by high concentrations of anti-TGF-beta suggesting that cell-associated TGF-beta1 was the primary effector of CD4(+)CD25(+)-mediated suppression. Here, we have reevaluated the role of TGF-beta1 in CD4(+)CD25(+)-mediated suppression. Neutralization of TGF-beta1 with either monoclonal antibody (mAb) or soluble TGF-betaRII-Fc did not reverse in vitro suppression mediated by resting or activated CD4(+)CD25(+) T cells. Responder T cells from Smad3(-/-) or dominant-negative TGF-beta type RII transgenic (DNRIITg) mice, that are both unresponsive to TGF-beta1-induced growth arrest, were as susceptible to CD4(+)CD25(+)-mediated suppression as T cells from wild-type mice. Furthermore, CD4(+)CD25(+) T cells from neonatal TGF-beta1(-/-) mice were as suppressive as CD4(+)CD25(+) from TGF-beta1(+/+) mice. Collectively, these results demonstrate that CD4(+)CD25(+) suppressor function can occur independently of TGF-beta1.
Figures

References
-
- Bach, J.F. 1995. Organ-specific autoimmunity. Immunol. Today. 16:353–355. - PubMed
-
- King, C., and N. Sarvetnick. 1997. Organ-specific autoimmunity. Curr. Opin. Immunol. 9:863–871. - PubMed
-
- Moller, G. 1996. Dominant immunological tolerance. Immunol. Rev. 149:1–17.
-
- Shevach, E.M. 2000. Regulatory T cells in autoimmmunity. Annu. Rev. Immunol. 18:423–449. - PubMed
-
- Sakaguchi, S. 2000. Regulatory T cells: key controllers of immunologic self-tolerance. Cell. 101:455–458. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
