

A phosphatidylinositol (4,5)-bisphosphate binding site within mu2-adaptin regulates clathrin-mediated endocytosis
- PMID: 12119359
- PMCID: PMC2173125
- DOI: 10.1083/jcb.200203103
A phosphatidylinositol (4,5)-bisphosphate binding site within mu2-adaptin regulates clathrin-mediated endocytosis
Abstract
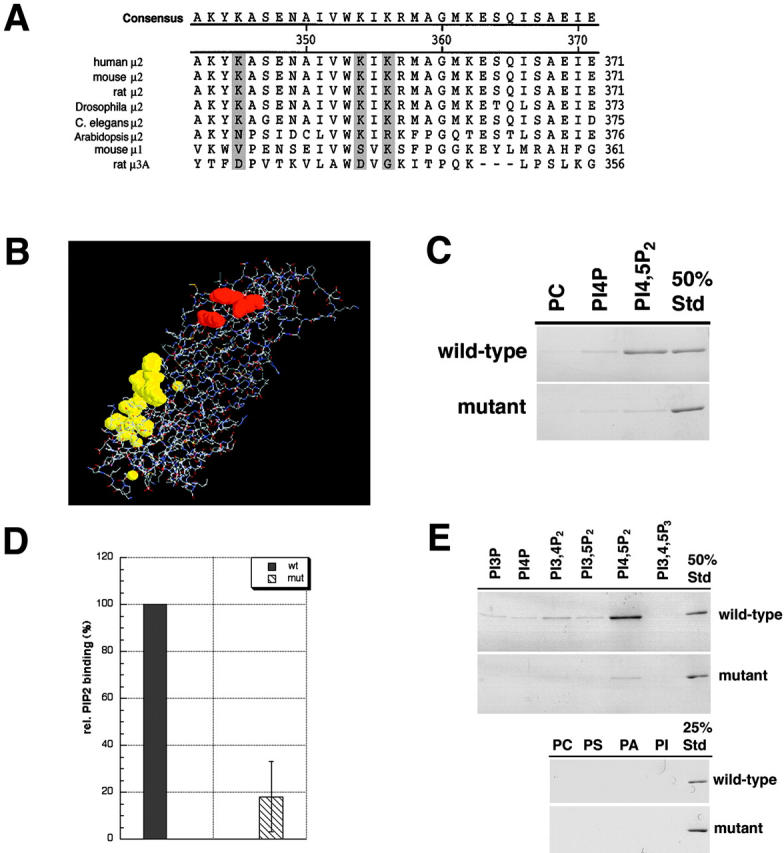
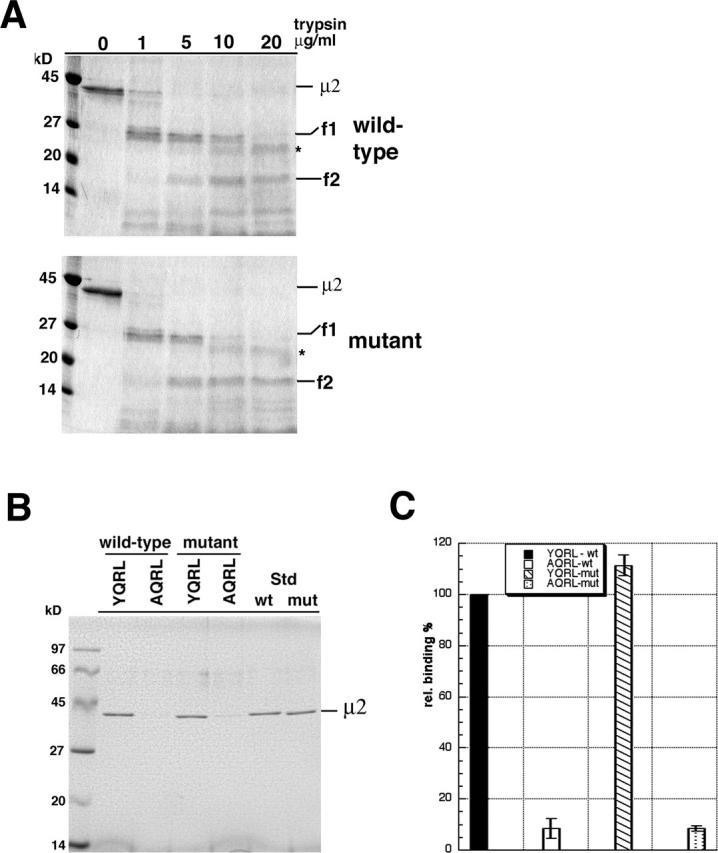
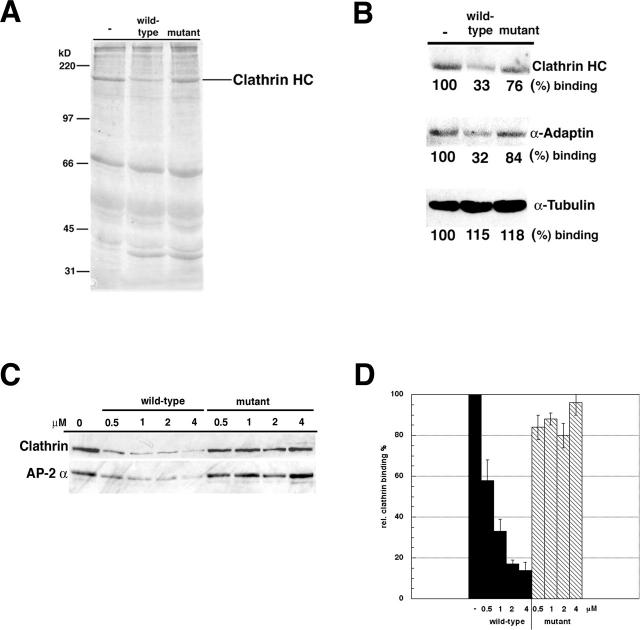
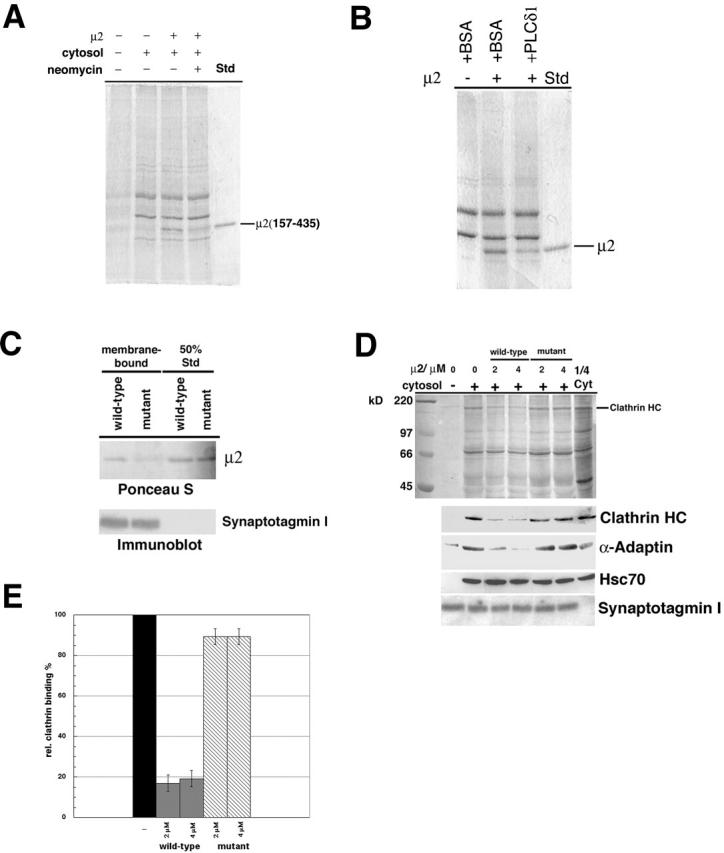
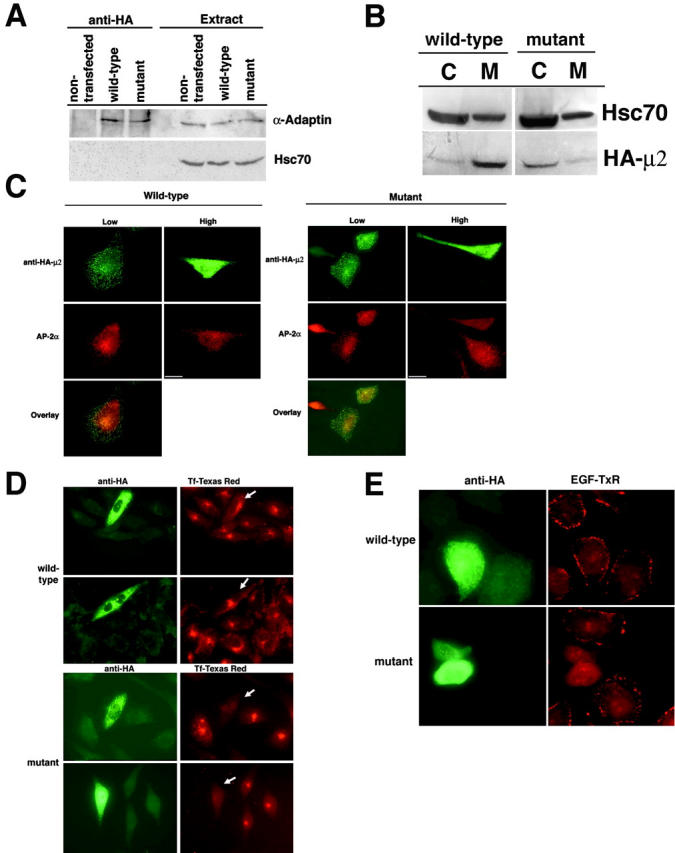
The clathrin adaptor complex AP-2 serves to coordinate clathrin-coated pit assembly with the sorting of transmembrane cargo proteins at the plasmalemma. How precisely AP-2 assembly and cargo protein recognition at sites of endocytosis are regulated has remained unclear, but recent evidence implicates phosphoinositides, in particular phosphatidylinositol (4,5)-bisphosphate (PI[4,5]P2), in these processes. Here we have identified and functionally characterized a conserved binding site for PI(4,5)P2 within mu2-adaptin, the medium chain of the clathrin adaptor complex AP-2. Mutant mu2 lacking a cluster of conserved lysine residues fails to bind PI(4,5)P2 and to compete the recruitment of native clathrin/AP-2 to PI(4,5)P2-containing liposomes or to presynaptic membranes. Moreover, we show that expression of mutant mu2 inhibits receptor-mediated endocytosis in living cells. We suggest that PI(4,5)P2 binding to mu2-adaptin regulates clathrin-mediated endocytosis and thereby may contribute to structurally linking cargo recognition to coat formation.
Figures
References
-
- Aguilar, R.C., H. Ohno, K.W. Roche, and J.S. Bonifacino. 1997. Functional domain mapping of the clathrin-associated adaptor medium chains mu1 and mu2. J. Biol. Chem. 272:27160–27166. - PubMed
-
- Arneson, L.S., J. Kunz, R.A. Anderson, and L.M. Traub. 1999. Coupled inositide phosphorylation and phospholipase D activation initiates clathrin-coat assembly on lysosomes. J. Biol. Chem. 274:17794–17805. - PubMed
-
- Brodin, L., P. Low, and O. Shupliakov. 2000. Sequential steps in clathrin-mediated synaptic vesicle endocytosis. Curr. Opin. Neurobiol. 10:312–320. - PubMed
-
- Bromann, P.A., E.E. Boetticher, and J.W. Lomasney. 1997. A single amino acid substitution in the pleckstrin homology domain of phospholipase C delta1 enhances the rate of substrate hydrolysis. J. Biol. Chem. 272:16240–16246. - PubMed
-
- Cremona, O., G. Di Paolo, M.R. Wenk, A. Luthi, W.T. Kim, K. Takei, L. Daniell, Y. Nemoto, S.B. Shears, R.A. Flavell, et al. 1999. Essential role of phosphoinositide metabolism in synaptic vesicle recycling. Cell. 99:179–188. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
