

Transcripts of Vp-1 homeologues are misspliced in modern wheat and ancestral species
- PMID: 12119408
- PMCID: PMC126648
- DOI: 10.1073/pnas.152318599
Transcripts of Vp-1 homeologues are misspliced in modern wheat and ancestral species
Abstract
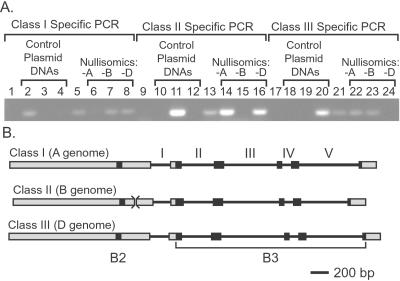
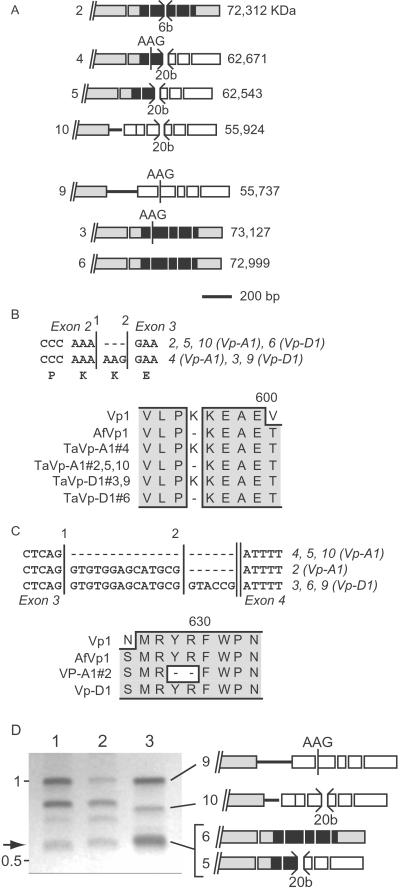
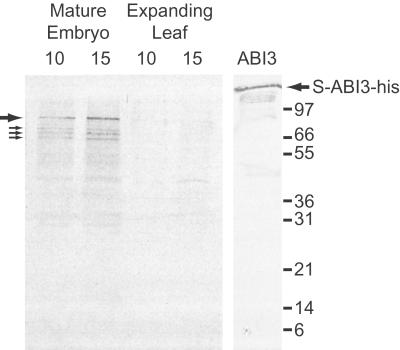
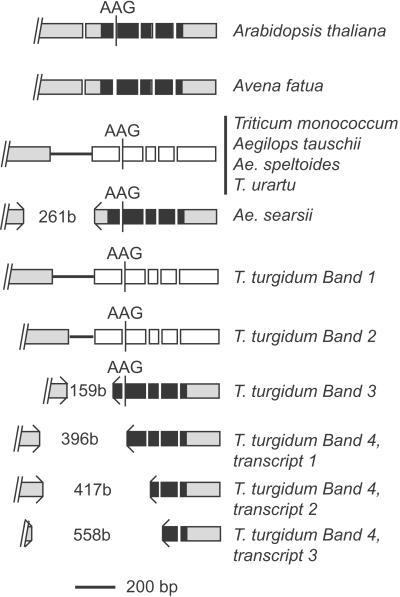
The maize (Zea mays) Viviparous 1 (Vp1) transcription factor has been shown previously to be a major regulator of seed development, simultaneously activating embryo maturation and repressing germination. Hexaploid bread wheat (Triticum aestivum) caryopses are characterized by relatively weak embryo dormancy and are susceptible to preharvest sprouting (PHS), a phenomenon that is phenotypically similar to the maize vp1 mutation. Analysis of Vp-1 transcript structure in wheat embryos during grain development showed that each homeologue produces cytoplasmic mRNAs of different sizes. The majority of transcripts are spliced incorrectly, contain insertions of intron sequences or deletions of coding region, and do not have the capacity to encode full-length proteins. Several VP-1-related lower molecular weight protein species were present in wheat embryo nuclei. Embryos of a closely related tetraploid species (Triticum turgidum) and ancestral diploids also contained misspliced Vp-1 transcripts that were structurally similar or identical to those found in modern hexaploid wheat, which suggests that compromised structure and expression of Vp-1 transcripts in modern wheat are inherited from ancestral species. Developing embryos from transgenic wheat grains expressing the Avena fatua Vp1 gene showed enhanced responsiveness to applied abscisic acid compared with the control. In addition, ripening ears of transgenic plants were less susceptible to PHS. Our results suggest that missplicing of wheat Vp-1 genes contributes to susceptibility to PHS in modern hexaploid wheat varieties and identifies a possible route to increase resistance to this environmentally triggered disorder.
Figures
References
-
- McCarty D R, Hattori T, Carson C B, Vasil V, Lazar M, Vasil I K. Cell. 1991;66:895–905. - PubMed
-
- Hoecker U, Vasil I K, McCarty D R. Genes Dev. 1995;9:2459–2469. - PubMed
-
- Hattori T, Terada T, Hamasuna S T. Plant Mol Biol. 1994;24:805–810. - PubMed
-
- Jones H D, Peters N C B, Holdsworth M J. Plant J. 1997;12:911–920. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
