

Graded contribution of the Gbeta gamma binding domains to GIRK channel activation
- PMID: 12124401
- PMCID: PMC125044
- DOI: 10.1073/pnas.162346199
Graded contribution of the Gbeta gamma binding domains to GIRK channel activation
Abstract
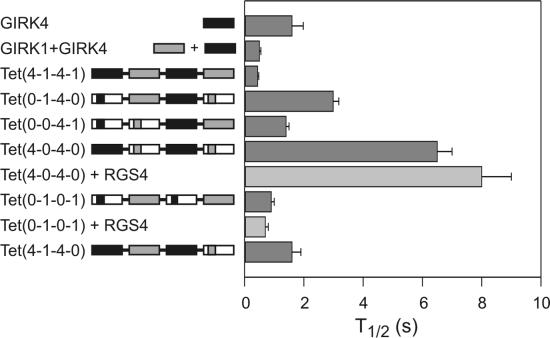
G protein coupled inwardly rectifying K(+) channels (GIRK/Kir3.x) are mainly activated by a direct interaction with Gbetagamma subunits, released upon the activation of inhibitory neurotransmitter receptors. Although Gbetagamma binding domains on all four subunits have been found, the relative contribution of each of these binding sites to channel gating has not yet been defined. It is also not known whether GIRK channels open once all Gbetagamma sites are occupied, or whether gating is a graded process. We used a tandem tetrameric approach to enable the selective elimination of specific Gbetagamma binding domains in the tetrameric context. Here, we show that tandem tetramers are fully operational. Tetramers with only one wild-type channel subunit showed receptor-independent high constitutive activity. The presence of two or three wild-type subunits reconstituted receptor activation gradually. Furthermore, a tetramer with no GIRK1 Gbetagamma binding domain displayed slower kinetics of activation. The slowdown in activation was found to be independent of regulator of G protein signaling or receptor coupling, but this slowdown could be reversed once only one Gbetagamma binding domain of GIRK1 was added. These results suggest that partial activation can occur under low Gbetagamma occupancy and that full activation can be accomplished by the interaction with three Gbetagamma binding subunits.
Figures



References
-
- Wickman K., Nemec, J., Gendler, S. J. & Clapham, D. E. (1998) Neuron 20, 103-114. - PubMed
-
- Luscher C., Jan, L. Y., Stoffel, M., Malenka, R. C. & Nicoll, R. A. (1997) Neuron 19, 687-695. - PubMed
-
- Reuveny E., Slesinger, P. A., Inglese, J., Morales, J. M., Iniguez-Lluhi, J. A., Lefkowitz, R. J., Bourne, H. R., Jan, Y. N. & Jan, L. Y. (1994) Nature (London) 370, 143-146. - PubMed
-
- Wickman K. D., Iniguez-Lluhl, J. A., Davenport, P. A., Taussig, R., Krapivinsky, G. B., Linder, M. E., Gilman, A. G. & Clapham, D. E. (1994) Nature (London) 368, 255-257. - PubMed
-
- Logothetis D. E., Kurachi, Y., Galper, J., Neer, E. J. & Clapham, D. E. (1987) Nature (London) 325, 321-326. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
