

Bovine leukemia virus SU protein interacts with zinc, and mutations within two interacting regions differently affect viral fusion and infectivity in vivo
- PMID: 12134000
- PMCID: PMC155115
- DOI: 10.1128/jvi.76.16.7956-7967.2002
Bovine leukemia virus SU protein interacts with zinc, and mutations within two interacting regions differently affect viral fusion and infectivity in vivo
Abstract
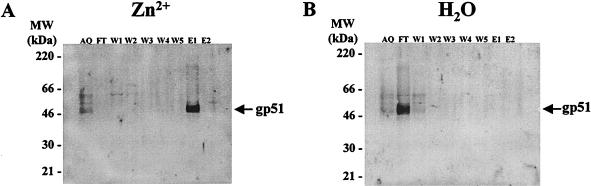
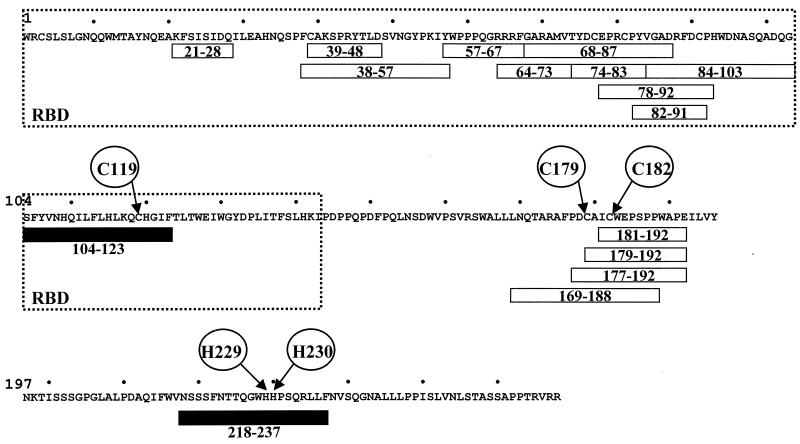
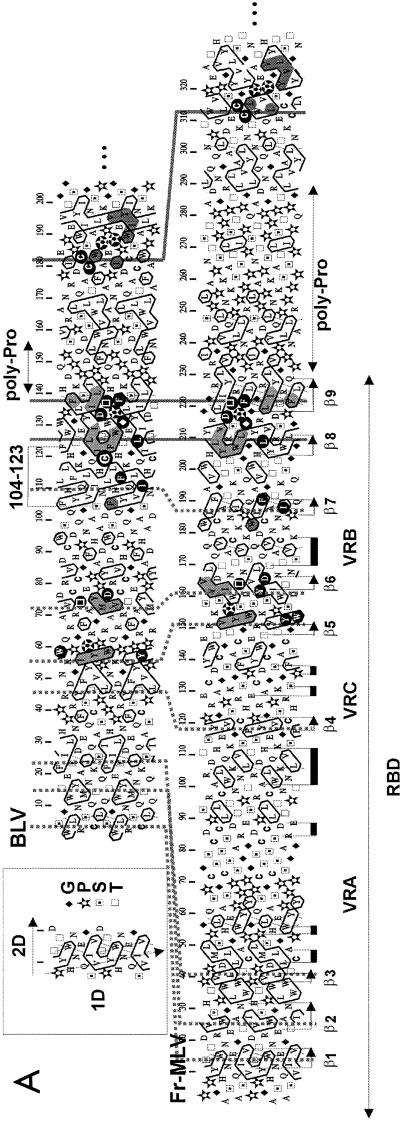
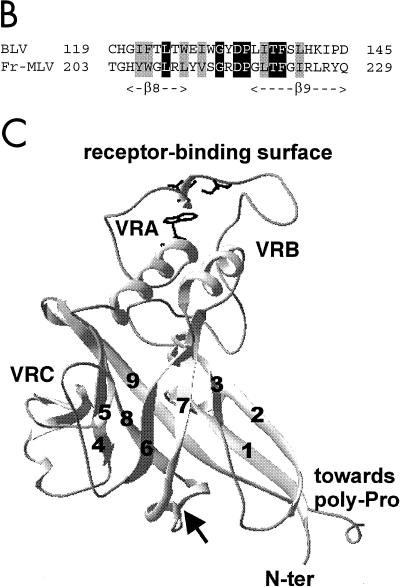
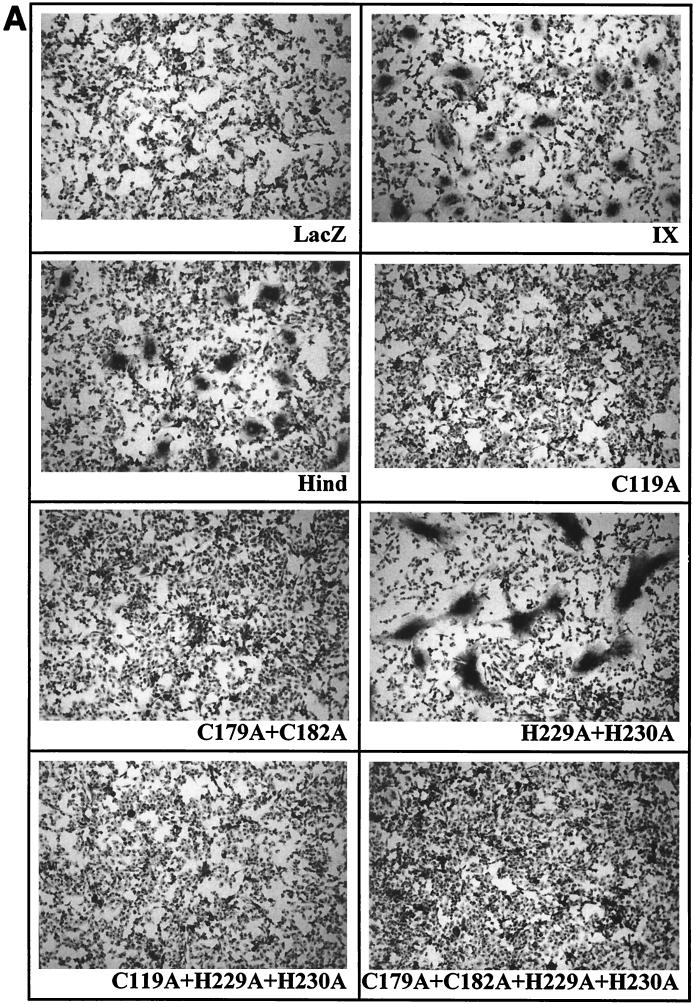
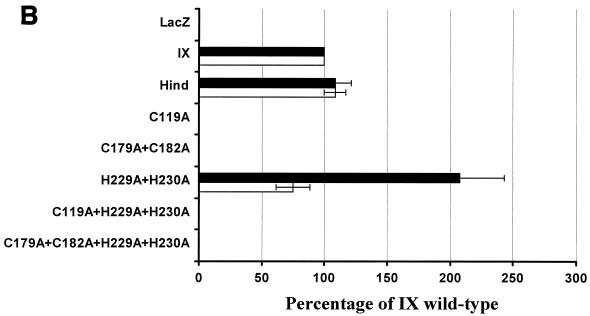
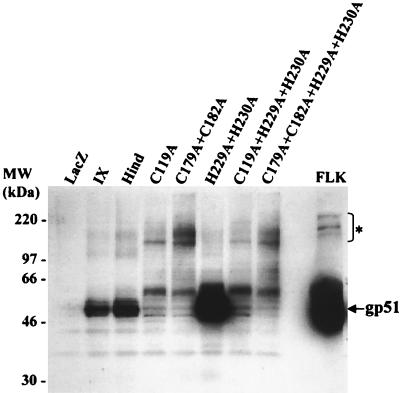
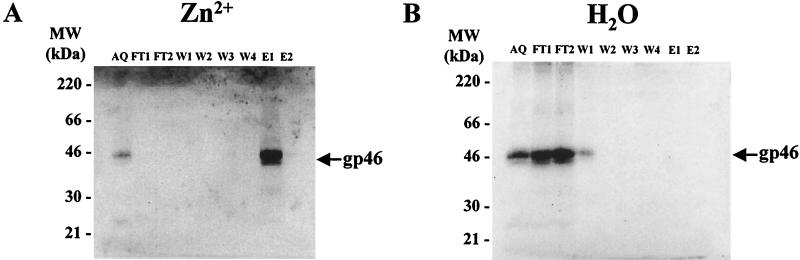
Bovine leukemia virus (BLV) and human T-cell lymphotropic virus type 1 (HTLV-1) belong to the genus of deltaretroviruses. Their entry into the host cell is supposed to be mediated by interactions of the extracellular (SU) envelope glycoproteins with cellular receptors. To gain insight into the mechanisms governing this process, we investigated the ability of SU proteins to interact with specific ligands. In particular, by affinity chromatography, we have shown that BLV SU protein specifically interacted with zinc ions. To identify the protein domains involved in binding, 16 peptides distributed along the sequence were tested. Two of them appeared to be able to interact with zinc. To unravel the role of these SU regions in the biology of the virus, mutations were introduced into the env gene of a BLV molecular clone in order to modify residues potentially interacting with zinc. The fusogenic capacity of envelope mutated within the first zinc-binding region (104 to 123) was completely abolished. Furthermore, the integrity of this domain was also required for in vivo infectivity. In contrast, mutations within the second zinc-binding region (218 to 237) did not hamper the fusogenic capacity; indeed, the syncytia were even larger. In sheep, mutations in region 218 to 237 did not alter infectivity or viral spread. Finally, we demonstrated that the envelope of the related HTLV-1 was also able to bind zinc. Interestingly, zinc ions were found to be associated with the receptor-binding domain (RBD) of Friend murine leukemia virus (Fr-MLV) SU glycoprotein, further supporting their relevance in SU structure. Based on the sequence similarities shared with the Fr-MLV RBD, whose three-dimensional structure has been experimentally determined, we located the BLV zinc-binding peptide 104-123 on the opposite side of the potential receptor-binding surface. This observation supports the hypothesis that zinc ions could mediate interactions of the SU RBD either with the C-terminal part of SU, thereby contributing to the SU structural integrity, or with a partner(s) different from the receptor.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
