

The herpes simplex virus type 1 US11 protein binds the coterminal UL12, UL13, and UL14 RNAs and regulates UL13 expression in vivo
- PMID: 12134014
- PMCID: PMC155164
- DOI: 10.1128/jvi.76.16.8090-8100.2002
The herpes simplex virus type 1 US11 protein binds the coterminal UL12, UL13, and UL14 RNAs and regulates UL13 expression in vivo
Abstract
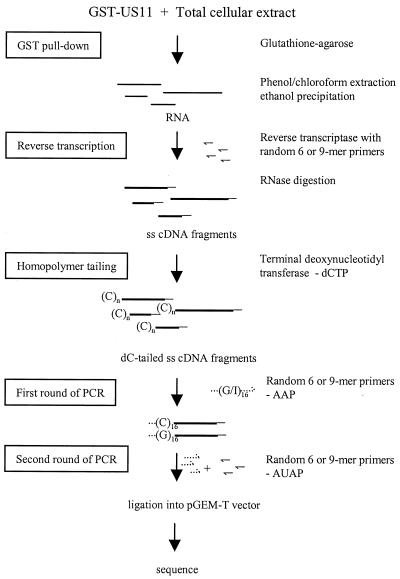
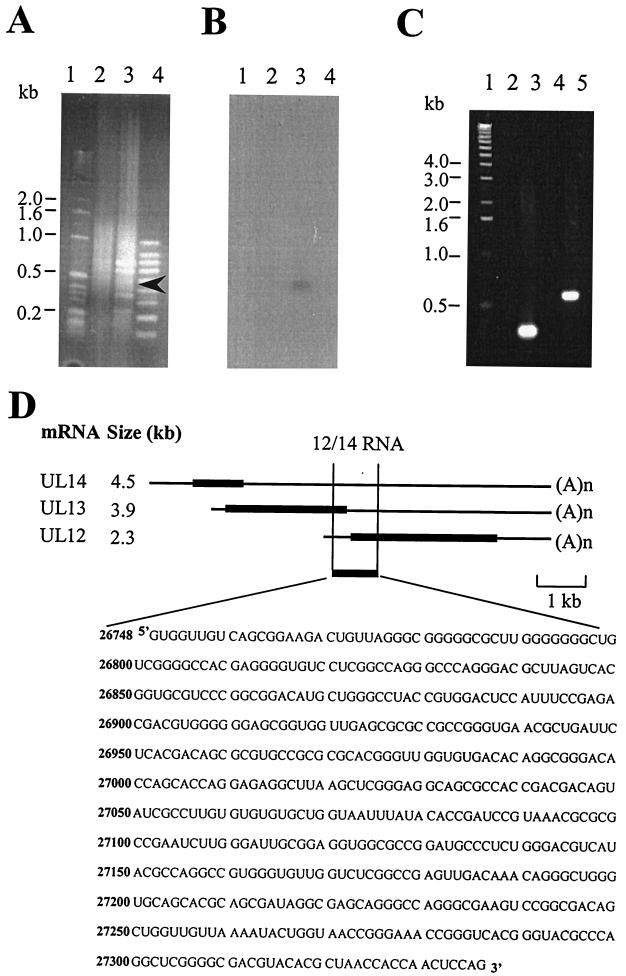
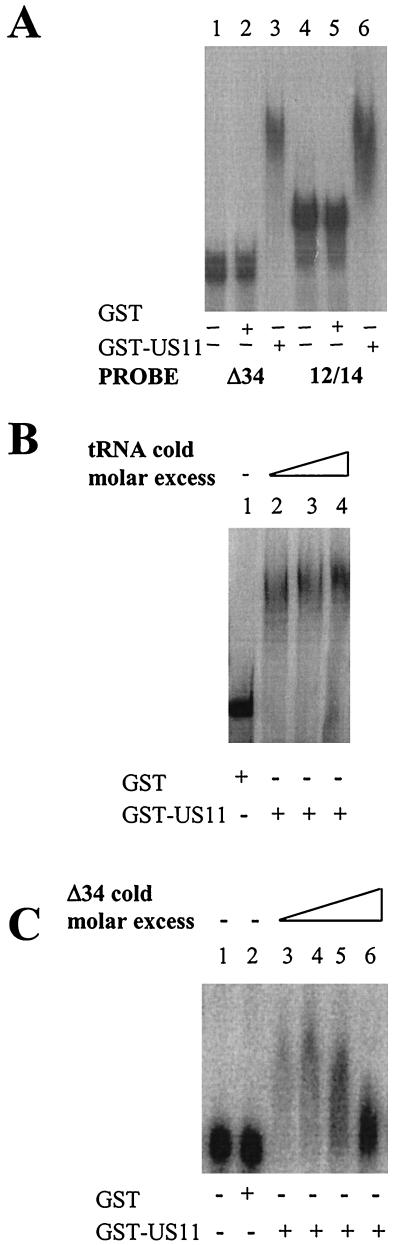
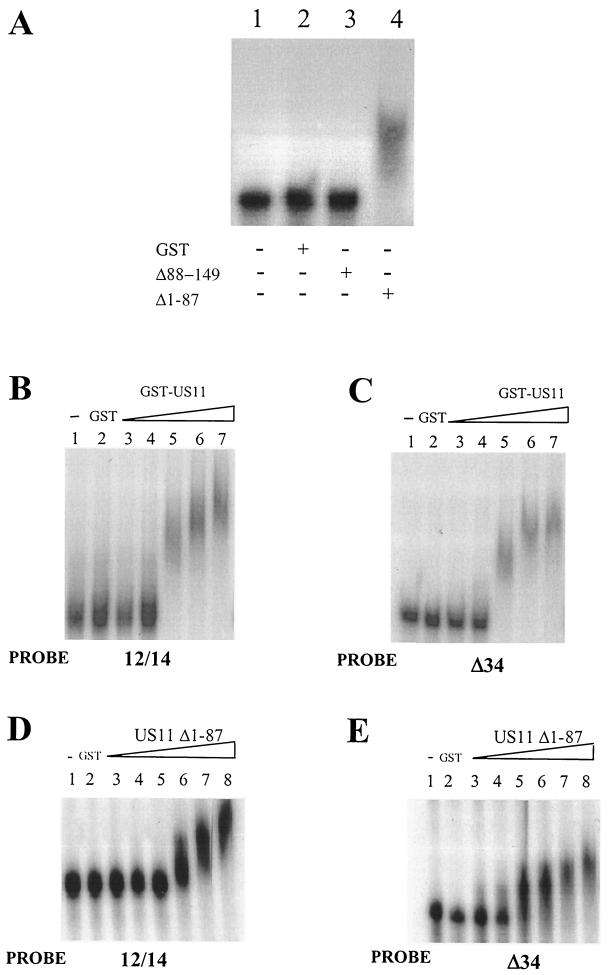
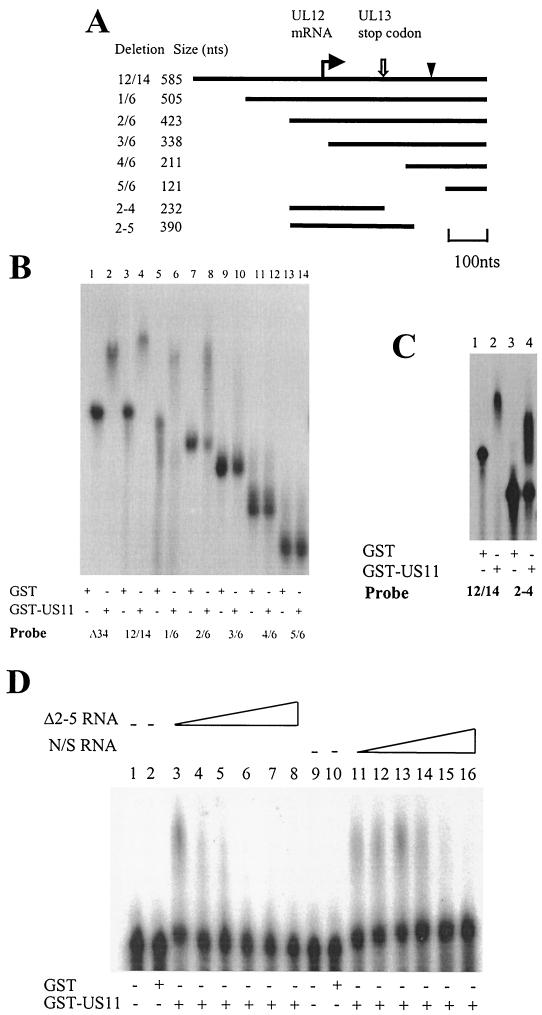
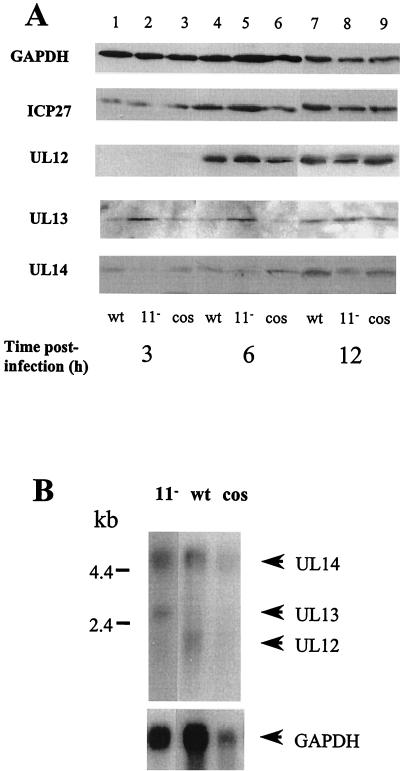
The US11 protein of herpes simplex virus type 1 (HSV-1) is a small, highly basic phosphoprotein expressed at late times during infection. US11 localizes to the nucleolus in infected cells, can associate with ribosomes, and has been shown to bind RNA. The RNA substrates of US11 identified thus far have no apparent role in the virus lytic cycle, so we set out to identify a novel, biologically relevant RNA substrate(s) for this protein in HSV-1-infected cells. We designed a reverse transcriptase PCR-based protocol that allowed specific selection of a 600-bp RNA binding partner for US11. This RNA sequence, designated 12/14, is present in the coterminal HSV-1 mRNAs UL12, UL13, and UL14. We show that the binding of US11 to 12/14 is sequence-specific and mediated by the C-terminal domain of the protein. To elucidate the role of US11 in the virus life cycle, we infected cells with wild-type virus, a cosmid-reconstructed US11 HSV-1 null mutant, and a cosmid-reconstructed wild-type virus and analyzed expression of UL12, -13, and -14 during a time course of infection. These experiments revealed that this interaction has biological activity; at early times of infection, US11 down-regulates UL13 protein kinase mRNA and protein.
Figures
References
-
- Brand, S. R., R. Kobayashi, and M. B. Matthews. 1997. The Tat protein of human immunodeficiency virus type 1 is a substrate and inhibitor of the interferon-induced, virally activated protein kinase, PKR. J. Biol. Chem. 272:8388-8395. - PubMed
-
- Cassady, K. A., M. Gross, and B. Roizman. 1998. The herpes simplex virus Us11 protein effectively compensates for the gamma134.5 gene if present before activation of protein kinase R by precluding its phosphorylation and that of the α subunit of eukaryotic translation initiation factor 2. J. Virol. 72:8620-8626. - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
