

Binding of human cytomegalovirus US2 to major histocompatibility complex class I and II proteins is not sufficient for their degradation
- PMID: 12134032
- PMCID: PMC155152
- DOI: 10.1128/jvi.76.16.8265-8275.2002
Binding of human cytomegalovirus US2 to major histocompatibility complex class I and II proteins is not sufficient for their degradation
Erratum in
- J Virol 2002 Dec;76(24):13126
Abstract
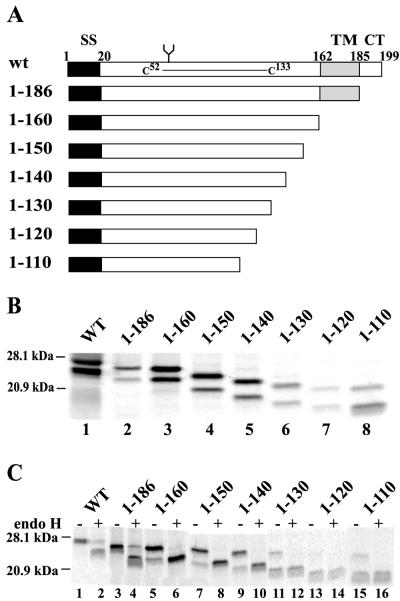
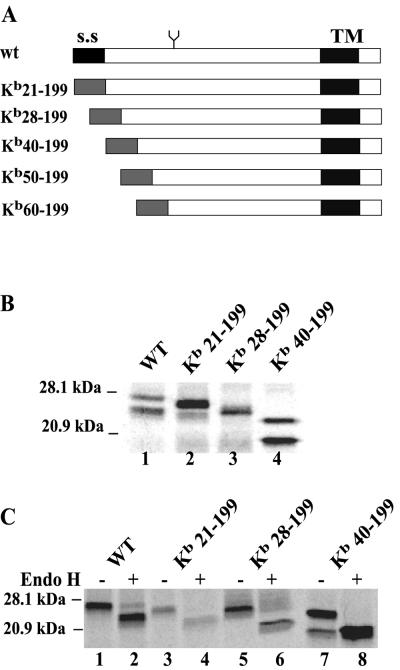
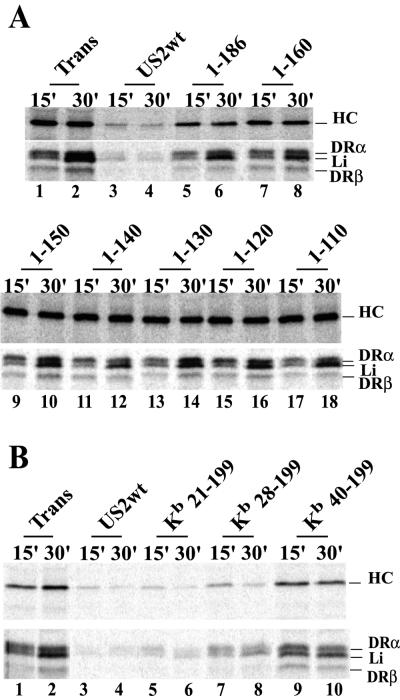
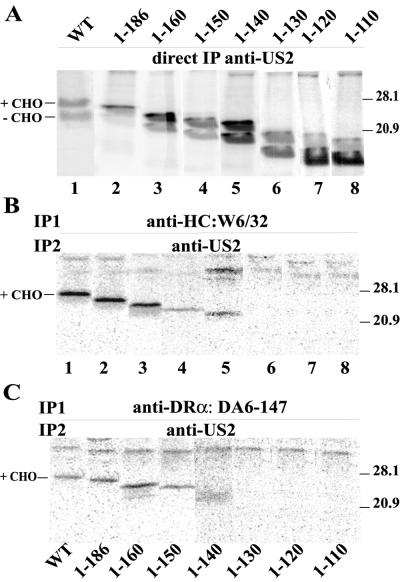
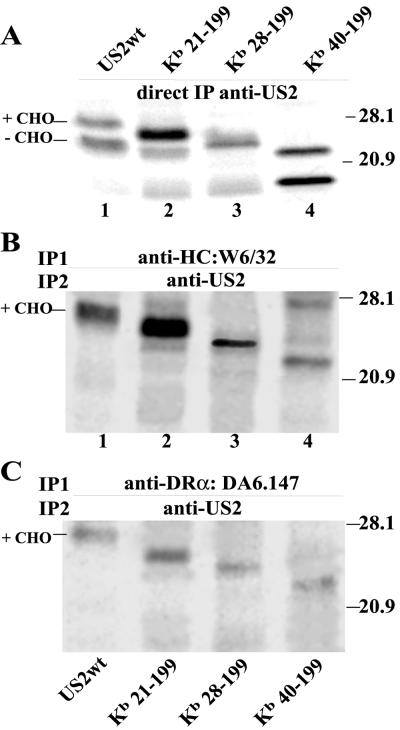
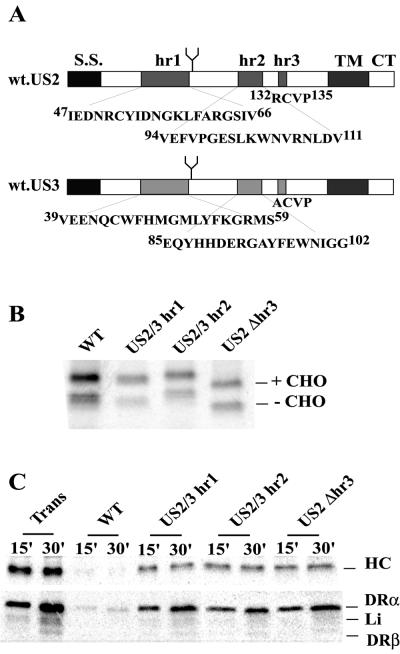
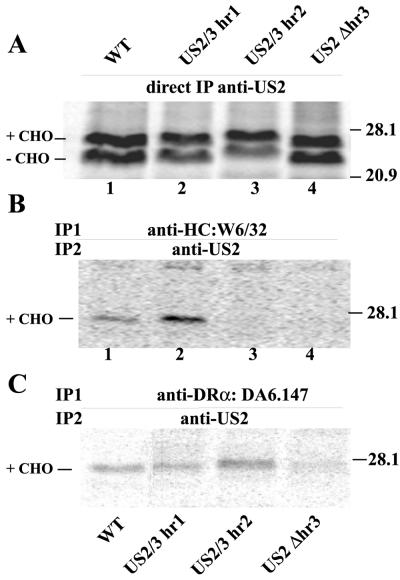
Human cytomegalovirus (HCMV) glycoprotein US2 causes degradation of major histocompatibility complex (MHC) class I heavy-chain (HC), class II DR-alpha and DM-alpha proteins, and HFE, a nonclassical MHC protein. In US2-expressing cells, MHC proteins present in the endoplasmic reticulum (ER) are degraded by cytosolic proteasomes. It appears that US2 binding triggers a normal cellular pathway by which misfolded or aberrant proteins are translocated from the ER to cytoplasmic proteasomes. To better understand how US2 binds MHC proteins and causes their degradation, we constructed a panel of US2 mutants. Mutants truncated from the N terminus as far as residue 40 or from the C terminus to amino acid 140 could bind to class I and class II proteins. Nevertheless, mutants lacking just the cytosolic tail (residues 187 to 199) were unable to cause degradation of both class I and II proteins. Chimeric proteins were constructed in which US2 sequences were replaced with homologous sequences from US3, an HCMV glycoprotein that can also bind to class I and II proteins. One of these US2/US3 chimeras bound to class II but not to class I, and a second bound class I HC better than wild-type US2. Therefore, US2 residues involved in the binding to MHC class I differ subtly from those involved in binding to class II proteins. Moreover, our results demonstrate that the binding of US2 to class I and II proteins is not sufficient to cause degradation of MHC proteins. The cytosolic tail of US2 and certain US2 lumenal sequences, which are not involved in binding to MHC proteins, are required for degradation. Our results are consistent with the hypothesis that US2 couples MHC proteins to components of the ER degradation pathway, enormously increasing the rate of degradation of MHC proteins.
Figures
Similar articles
-
Human cytomegalovirus US3 chimeras containing US2 cytosolic residues acquire major histocompatibility class I and II protein degradation properties.J Virol. 2003 Apr;77(8):4731-8. doi: 10.1128/jvi.77.8.4731-4738.2003. J Virol. 2003. PMID: 12663780 Free PMC article.
-
Inhibition of the MHC class II antigen presentation pathway by human cytomegalovirus.Curr Top Microbiol Immunol. 2002;269:101-15. doi: 10.1007/978-3-642-59421-2_7. Curr Top Microbiol Immunol. 2002. PMID: 12224504 Review.
-
Human cytomegalovirus US2 causes similar effects on both major histocompatibility complex class I and II proteins in epithelial and glial cells.J Virol. 2003 Sep;77(17):9287-94. doi: 10.1128/jvi.77.17.9287-9294.2003. J Virol. 2003. PMID: 12915544 Free PMC article.
-
Human cytomegalovirus-encoded US2 differentially affects surface expression of MHC class I locus products and targets membrane-bound, but not soluble HLA-G1 for degradation.J Immunol. 2003 Dec 15;171(12):6757-65. doi: 10.4049/jimmunol.171.12.6757. J Immunol. 2003. PMID: 14662880
-
The HCMV gene products US2 and US11 target MHC class I molecules for degradation in the cytosol.Curr Top Microbiol Immunol. 2002;269:37-55. doi: 10.1007/978-3-642-59421-2_3. Curr Top Microbiol Immunol. 2002. PMID: 12224515 Review.
Cited by
-
A slowly cleaved viral signal peptide acts as a protein-integral immune evasion domain.Nat Commun. 2021 Apr 6;12(1):2061. doi: 10.1038/s41467-021-21983-x. Nat Commun. 2021. PMID: 33824318 Free PMC article.
-
A novel viral transcript with homology to human interleukin-10 is expressed during latent human cytomegalovirus infection.J Virol. 2004 Feb;78(3):1440-7. doi: 10.1128/jvi.78.3.1440-1447.2004. J Virol. 2004. PMID: 14722299 Free PMC article.
-
Human cytomegalovirus evades antibody-mediated immunity through endoplasmic reticulum-associated degradation of the FcRn receptor.Nat Commun. 2019 Jul 9;10(1):3020. doi: 10.1038/s41467-019-10865-y. Nat Commun. 2019. PMID: 31289263 Free PMC article.
-
The correlation between prognosis of HCMV infection and zinc in mature women.J Huazhong Univ Sci Technolog Med Sci. 2004;24(6):628-9. doi: 10.1007/BF02911376. J Huazhong Univ Sci Technolog Med Sci. 2004. PMID: 15791862
-
Manipulation of iron to determine survival: competition between host and pathogen.Immunol Res. 2004;30(1):15-28. doi: 10.1385/IR:30:1:015. Immunol Res. 2004. PMID: 15258308 Review.
References
-
- Ahn, K., A. Gruhler, B. Galocha, T. R. Jones, E. J. Wiertz, H. L. Ploegh, P. A. Peterson, Y. Yang, and K. Fruh. 1997. The ER-luminal domain of the HCMV glycoprotein US6 inhibits peptide translocation by TAP. Immunity 6:613-621. - PubMed
-
- Beersma, M. F., M. J. Bijlmakers, and H. L. Ploegh. 1993. Human cytomegalovirus downregulates HLA class I expression by reducing the stability of class I H chains. J. Immunol. 151:4455-4464. - PubMed
-
- Ben-Arieh, S. V., B. Zimerman, N. I. Smorodinsky, M. Yaacubovicz, C. Schechter, I. Bacik, J. Gibbs, J. R. Bennink, J. W. Yewdell, J. E. Coligan, H. Firat, F. Lemonnier, and R. Ehrlich. 2001. Human cytomegalovirus protein US2 interferes with the expression of human HFE, a nonclassical class I major histocompatibility complex molecule that regulates iron homeostasis. J. Virol. 75:10557-10562. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
