

Disruption of centrosome structure, chromosome segregation, and cytokinesis by misexpression of human Cdc14A phosphatase
- PMID: 12134069
- PMCID: PMC117313
- DOI: 10.1091/mbc.01-11-0535
Disruption of centrosome structure, chromosome segregation, and cytokinesis by misexpression of human Cdc14A phosphatase
Abstract
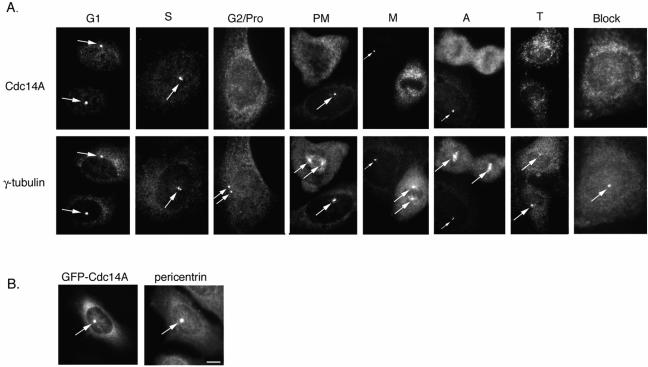
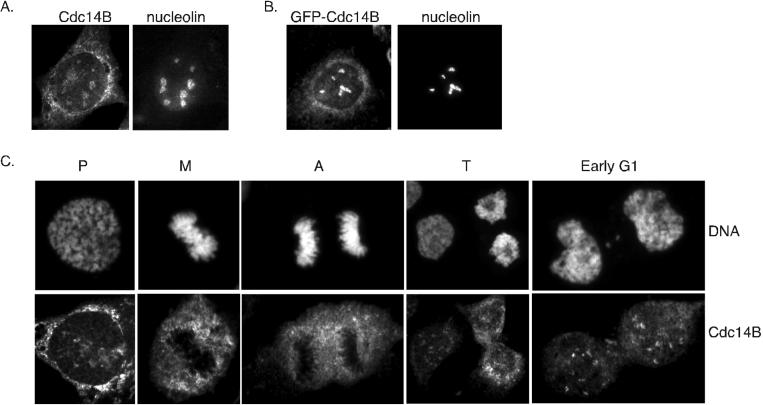
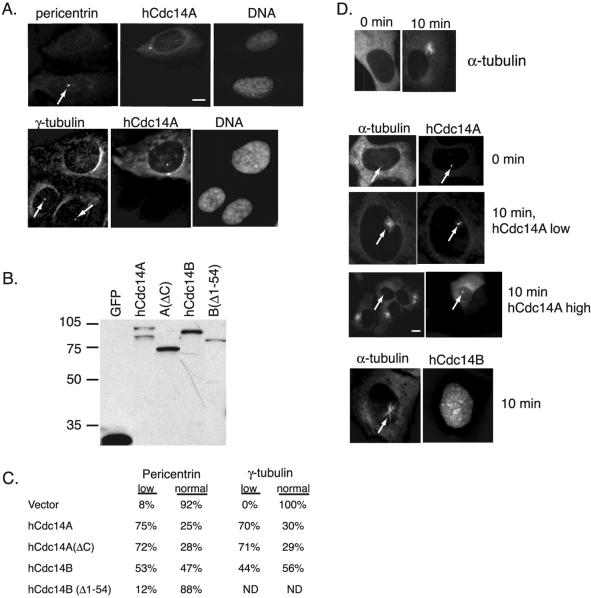
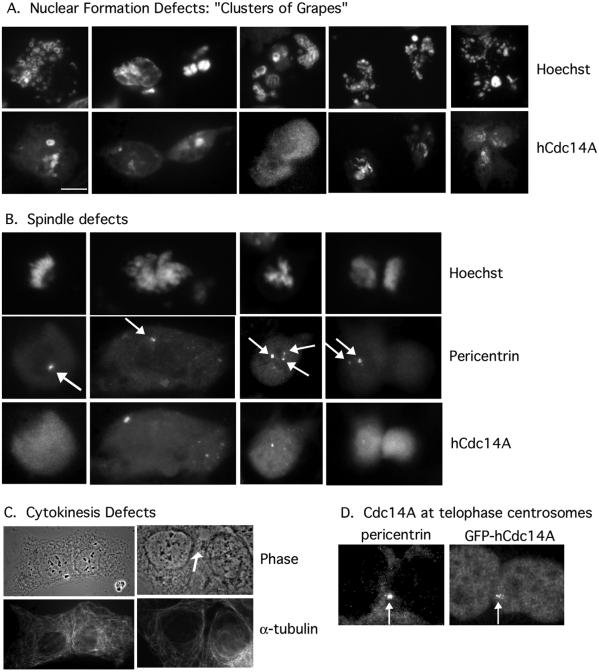
In budding yeast, the Cdc14p phosphatase activates mitotic exit by dephosphorylation of specific cyclin-dependent kinase (Cdk) substrates and seems to be regulated by sequestration in the nucleolus until its release in mitosis. Herein, we have analyzed the two human homologs of Cdc14p, hCdc14A and hCdc14B. We demonstrate that the human Cdc14A phosphatase is selective for Cdk substrates in vitro and that although the protein abundance and intrinsic phosphatase activity of hCdc14A and B vary modestly during the cell cycle, their localization is cell cycle regulated. hCdc14A dynamically localizes to interphase but not mitotic centrosomes, and hCdc14B localizes to the interphase nucleolus. These distinct patterns of localization suggest that each isoform of human Cdc14 likely regulates separate cell cycle events. In addition, hCdc14A overexpression induces the loss of the pericentriolar markers pericentrin and gamma-tubulin from centrosomes. Overproduction of hCdc14A also causes mitotic spindle and chromosome segregation defects, defective karyokinesis, and a failure to complete cytokinesis. Thus, the hCdc14A phosphatase appears to play a role in the regulation of the centrosome cycle, mitosis, and cytokinesis, thereby influencing chromosome partitioning and genomic stability in human cells.
Figures

Similar articles
-
Deregulated human Cdc14A phosphatase disrupts centrosome separation and chromosome segregation.Nat Cell Biol. 2002 Apr;4(4):317-22. doi: 10.1038/ncb777. Nat Cell Biol. 2002. PMID: 11901424
-
HCdc14A is involved in cell cycle regulation of human brain vascular endothelial cells following injury induced by high glucose, free fatty acids and hypoxia.Cell Signal. 2015 Jan;27(1):47-60. doi: 10.1016/j.cellsig.2014.10.003. Epub 2014 Oct 16. Cell Signal. 2015. PMID: 25463242
-
Functional homology among human and fission yeast Cdc14 phosphatases.J Biol Chem. 2005 Aug 12;280(32):29144-50. doi: 10.1074/jbc.M413328200. Epub 2005 May 23. J Biol Chem. 2005. PMID: 15911625
-
Dynamic changes in nuclear architecture during mitosis: on the role of protein phosphorylation in spindle assembly and chromosome segregation.Exp Cell Res. 1996 Dec 15;229(2):174-80. doi: 10.1006/excr.1996.0356. Exp Cell Res. 1996. PMID: 8986594 Review.
-
Mitotic phosphatases: no longer silent partners.Curr Opin Cell Biol. 2006 Dec;18(6):623-31. doi: 10.1016/j.ceb.2006.09.001. Epub 2006 Oct 9. Curr Opin Cell Biol. 2006. PMID: 17030123 Review.
Cited by
-
The structure of the cell cycle protein Cdc14 reveals a proline-directed protein phosphatase.EMBO J. 2003 Jul 15;22(14):3524-35. doi: 10.1093/emboj/cdg348. EMBO J. 2003. PMID: 12853468 Free PMC article.
-
The Clp1/Cdc14 phosphatase contributes to the robustness of cytokinesis by association with anillin-related Mid1.J Cell Biol. 2008 Apr 7;181(1):79-88. doi: 10.1083/jcb.200709060. Epub 2008 Mar 31. J Cell Biol. 2008. PMID: 18378776 Free PMC article.
-
Regulation of APC/C-Cdh1 and its function in neuronal survival.Mol Neurobiol. 2012 Dec;46(3):547-54. doi: 10.1007/s12035-012-8309-2. Epub 2012 Jul 27. Mol Neurobiol. 2012. PMID: 22836916 Free PMC article. Review.
-
The dual-specificity phosphatase CDC14B bundles and stabilizes microtubules.Mol Cell Biol. 2005 Jun;25(11):4541-51. doi: 10.1128/MCB.25.11.4541-4551.2005. Mol Cell Biol. 2005. PMID: 15899858 Free PMC article.
-
Dynamic regulation of the PR-Set7 histone methyltransferase is required for normal cell cycle progression.Genes Dev. 2010 Nov 15;24(22):2531-42. doi: 10.1101/gad.1984210. Epub 2010 Oct 21. Genes Dev. 2010. PMID: 20966048 Free PMC article.
References
-
- Balasubramanian MK, McCollum D, Surana U. Tying the knot: linking cytokinesis to the nuclear cycle. J Cell Sci. 2000;113:1503–1513. - PubMed
-
- Brachmann CB, Sherman JM, Devine SE, Cameron EE, Pillus L, Boeke JD. The SIR2 gene family, conserved from bacteria to humans, functions in silencing, cell cycle progression, and chromosome stability. Genes Dev. 1995;9:2888–2902. - PubMed
-
- Carrano AC, Eytan E, Hershko A, Pagano M. SKP2 is required for ubiquitin-mediated degradation of the CDK inhibitor p27. Nat Cell Biol. 1999;1:193–199. - PubMed
-
- Chang P, Stearns T. δ-Tubulin and ε-tubulin: two new human centrosomal tubulins reveal new aspects of centrosome structure and function. Nat Cell Biol. 2000;2:30–35. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
