

Alternate exon insertion controls selective ubiquitination and degradation of different AUF1 protein isoforms
- PMID: 12136087
- PMCID: PMC135764
- DOI: 10.1093/nar/gkf444
Alternate exon insertion controls selective ubiquitination and degradation of different AUF1 protein isoforms
Abstract
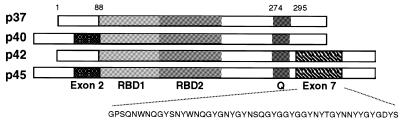
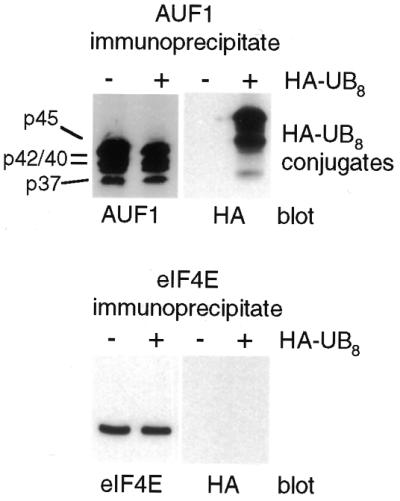
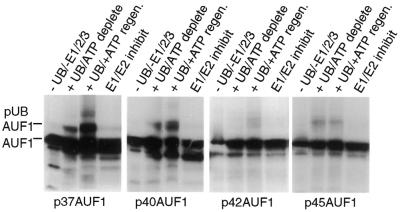
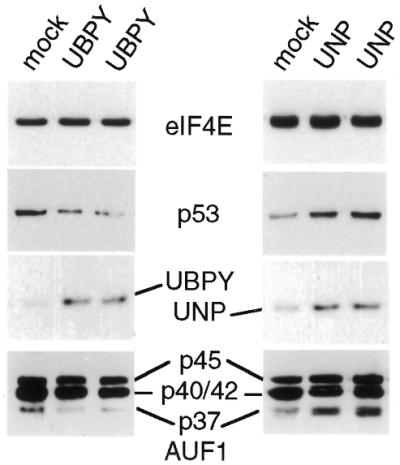
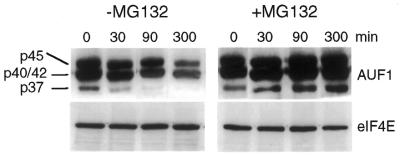
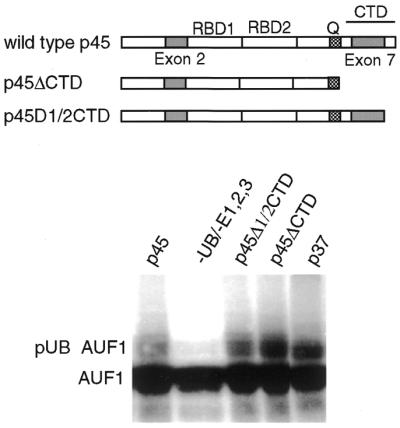
The A+U-rich element (ARE) in the 3' non-coding region (3' NCR) of short-lived cytokine mRNAs binds several regulatory proteins, including hnRNP D/AUF1, which comprises four isoforms of 37, 40, 42 and 45 kDa. ARE-mRNA degradation involves ubiquitin-proteasome activity, and one or more AUF1 proteins are thought to be ubiquitinated. Here we have characterized the mechanism for differential ubiquitination and degradation of the different AUF1 protein isoforms. We demonstrate in an in vitro ubiquitination system that the p37, followed by the p40 protein, are strongly ubiquitinated, whereas the p42 and p45 forms are not. Over expression in cells of enzymes that control the ubiquitin cycle were found to control p37 and p40 AUF1 protein levels through ubiquitination and proteasome activity, but not p42 and p45 forms. The p42 and p45 AUF1 proteins share a C-terminal exon 7 that is not found in the p37/p40 isoforms. Our studies show that exon 7 blocks ubiquitination and rapid degradation of AUF1 proteins, whereas its deletion permits ubiquitination to occur and promotes rapid turnover of AUF1 proteins. Thus, the stabilities of AUF1 isoforms are differentially controlled by insertion of an alternate exon that regulates ubiquitin targeting activity.
Figures
Similar articles
-
Selective degradation of AU-rich mRNAs promoted by the p37 AUF1 protein isoform.Mol Cell Biol. 2003 Sep;23(18):6685-93. doi: 10.1128/MCB.23.18.6685-6693.2003. Mol Cell Biol. 2003. PMID: 12944492 Free PMC article.
-
14-3-3sigma is a p37 AUF1-binding protein that facilitates AUF1 transport and AU-rich mRNA decay.EMBO J. 2006 Aug 23;25(16):3823-31. doi: 10.1038/sj.emboj.7601264. Epub 2006 Aug 10. EMBO J. 2006. PMID: 16902409 Free PMC article.
-
A nuclear matrix-associated factor, SAF-B, interacts with specific isoforms of AUF1/hnRNP D.Arch Biochem Biophys. 2000 Aug 15;380(2):228-36. doi: 10.1006/abbi.2000.1938. Arch Biochem Biophys. 2000. PMID: 10933876
-
Physiological networks and disease functions of RNA-binding protein AUF1.Wiley Interdiscip Rev RNA. 2014 Jul-Aug;5(4):549-64. doi: 10.1002/wrna.1230. Epub 2014 Mar 28. Wiley Interdiscip Rev RNA. 2014. PMID: 24687816 Review.
-
Modulation of neoplastic gene regulatory pathways by the RNA-binding factor AUF1.Front Biosci (Landmark Ed). 2011 Jun 1;16(6):2307-25. doi: 10.2741/3855. Front Biosci (Landmark Ed). 2011. PMID: 21622178 Free PMC article. Review.
Cited by
-
Post-transcriptional regulatory networks in immunity.Immunol Rev. 2013 May;253(1):253-72. doi: 10.1111/imr.12051. Immunol Rev. 2013. PMID: 23550651 Free PMC article. Review.
-
Concurrent binding and modifications of AUF1 and HuR mediate the pH-responsive stabilization of phosphoenolpyruvate carboxykinase mRNA in kidney cells.Am J Physiol Renal Physiol. 2012 Dec 1;303(11):F1545-54. doi: 10.1152/ajprenal.00400.2012. Epub 2012 Sep 26. Am J Physiol Renal Physiol. 2012. PMID: 23019227 Free PMC article.
-
Reduction of c-Fos via Overexpression of miR-34a Results in Enhancement of TNF- Production by LPS in Neutrophils from Myelodysplastic Syndrome Patients.PLoS One. 2016 Aug 11;11(8):e0158527. doi: 10.1371/journal.pone.0158527. eCollection 2016. PLoS One. 2016. PMID: 27513856 Free PMC article.
-
Selective degradation of AU-rich mRNAs promoted by the p37 AUF1 protein isoform.Mol Cell Biol. 2003 Sep;23(18):6685-93. doi: 10.1128/MCB.23.18.6685-6693.2003. Mol Cell Biol. 2003. PMID: 12944492 Free PMC article.
-
Differential expression of RNA-binding proteins in bronchial epithelium of stable COPD patients.Int J Chron Obstruct Pulmon Dis. 2018 Oct 12;13:3173-3190. doi: 10.2147/COPD.S166284. eCollection 2018. Int J Chron Obstruct Pulmon Dis. 2018. PMID: 30349226 Free PMC article.
References
-
- Mitchell P., and Tollervey,D. (2000) mRNA stability in eukaryotes. Curr. Opin. Genet. Dev., 10, 193–198. - PubMed
-
- Guhaniyogi J., and Brewer,G. (2001) Regulation of mRNA stability in mammalian cells. Gene, 265, 11–23. - PubMed
-
- Wilusz C.J., Wormington,M. and Peltz,S.W. (2001) The cap-to-tail guide to mRNA turnover. Nature Rev. Mol. Cell. Biol., 2, 237–246. - PubMed
-
- Chen C.-Y.A., and Shyu,A.-B. (1995) AU-rich elements: characterization and importance in mRNA degradation. Trends Biochem. Sci., 20, 465–470. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
