

Identification of Sox-2 regulatory region which is under the control of Oct-3/4-Sox-2 complex
- PMID: 12136102
- PMCID: PMC135755
- DOI: 10.1093/nar/gkf435
Identification of Sox-2 regulatory region which is under the control of Oct-3/4-Sox-2 complex
Abstract
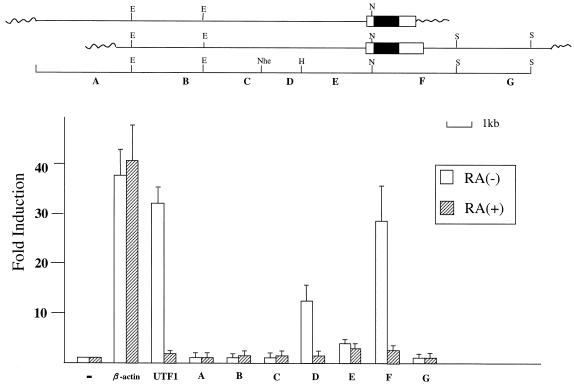
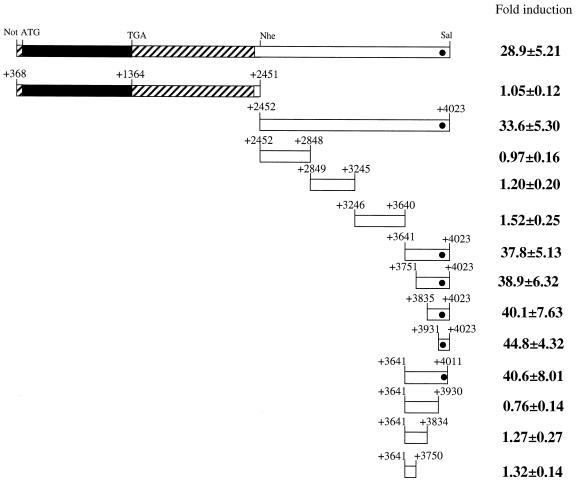
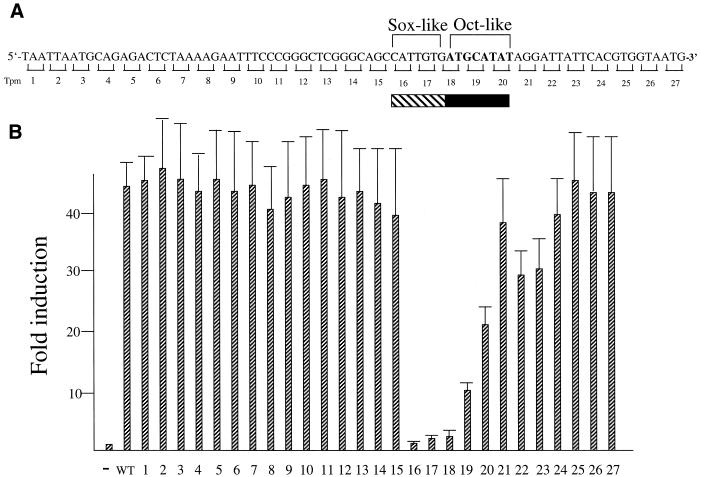
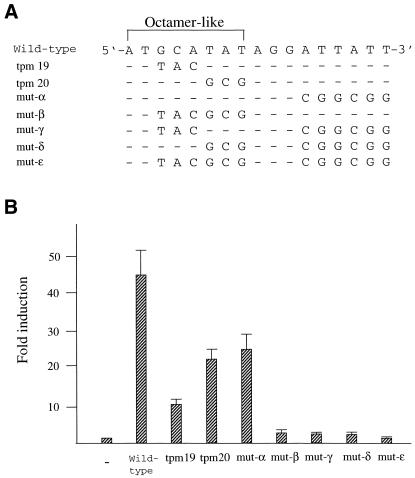
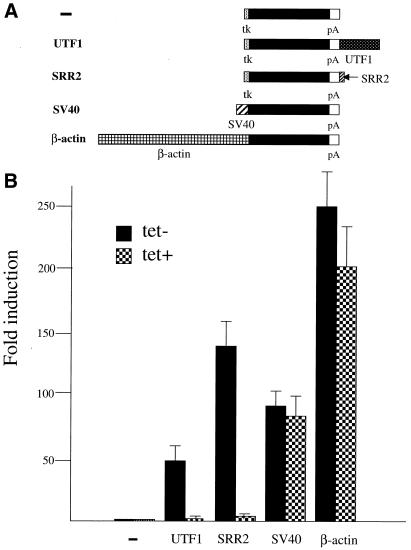
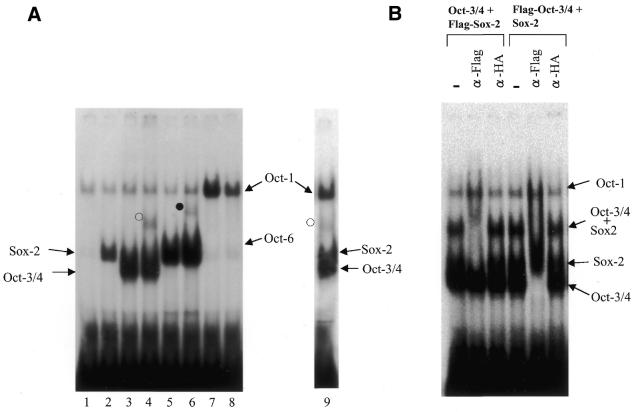
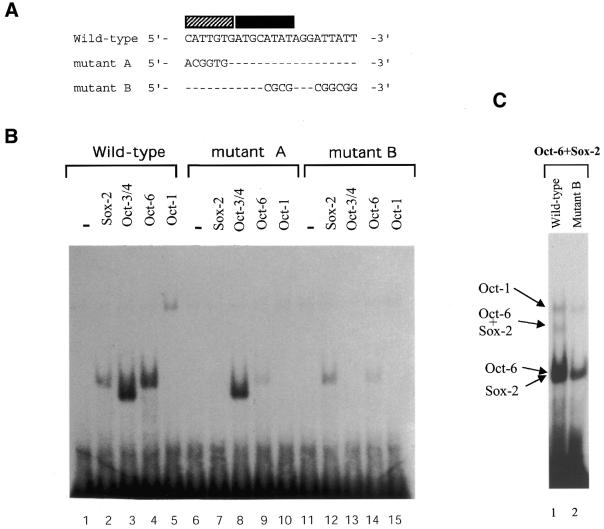
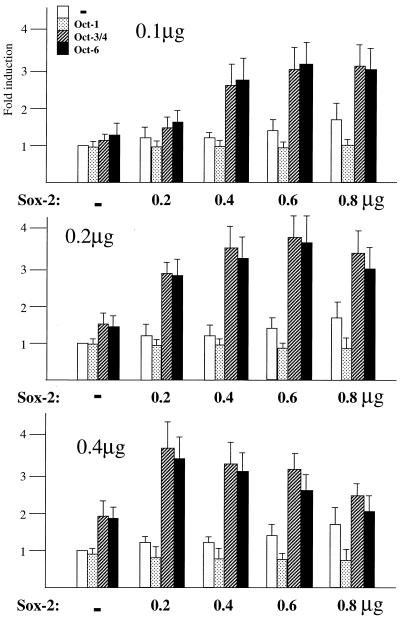
Sox-2 is a transcriptional cofactor expressed in embryonic stem (ES) cells as well as in neuronal cells. It has been demonstrated that Sox-2 plays an important role in supporting gene expression in ES cells, especially by forming a complex with embryonic Octamer factor, Oct-3/4. Here, we have analyzed the regulatory regions of the Sox-2 gene and identified two enhancers which stimulate transcription in ES cells as well as in embryonal carcinoma cells. These regulatory regions, which we termed Sox regulatory regions (SRR) 1 and 2, exert their function specifically when cells are in an undifferentiated state. Interestingly, like the regulatory elements of FGF-4 and UTF1 genes, combinatorial action of Octamer and Sox-2 binding sites support the SRR2 activity. However, biochemical analyses reveal that, due to the unique sequence and/or its organization, the SRR2 bears distinct characteristics from those of FGF-4 and UTF1 regulatory elements. That is, unlike the FGF-4 gene enhancer, the SRR2 precludes the binding of the Oct-1-Sox-2 complex. The difference between the SRR2 and UTF1 regulatory element is in the ability of SRR2 to recruit the Oct-6-Sox-2 complex as well as the Oct-3/4-Sox-2 complex. Co-transfection analyses confirm that both complexes are able to stimulate transcription through the SRR2 element.
Figures

Similar articles
-
The gene for the embryonic stem cell coactivator UTF1 carries a regulatory element which selectively interacts with a complex composed of Oct-3/4 and Sox-2.Mol Cell Biol. 1999 Aug;19(8):5453-65. doi: 10.1128/MCB.19.8.5453. Mol Cell Biol. 1999. PMID: 10409735 Free PMC article.
-
Oct-3/4 maintains the proliferative embryonic stem cell state via specific binding to a variant octamer sequence in the regulatory region of the UTF1 locus.Mol Cell Biol. 2005 Jun;25(12):5084-94. doi: 10.1128/MCB.25.12.5084-5094.2005. Mol Cell Biol. 2005. PMID: 15923625 Free PMC article.
-
Developmental-specific activity of the FGF-4 enhancer requires the synergistic action of Sox2 and Oct-3.Genes Dev. 1995 Nov 1;9(21):2635-45. doi: 10.1101/gad.9.21.2635. Genes Dev. 1995. PMID: 7590241
-
Pairing SOX off: with partners in the regulation of embryonic development.Trends Genet. 2000 Apr;16(4):182-7. doi: 10.1016/s0168-9525(99)01955-1. Trends Genet. 2000. PMID: 10729834 Review.
-
Evolution of non-coding regulatory sequences involved in the developmental process: reflection of differential employment of paralogous genes as highlighted by Sox2 and group B1 Sox genes.Proc Jpn Acad Ser B Phys Biol Sci. 2009;85(2):55-68. doi: 10.2183/pjab.85.55. Proc Jpn Acad Ser B Phys Biol Sci. 2009. PMID: 19212098 Free PMC article. Review.
Cited by
-
Sox2: regulation of expression and contribution to brain tumors.CNS Oncol. 2016 Jul;5(3):159-73. doi: 10.2217/cns-2016-0001. Epub 2016 May 27. CNS Oncol. 2016. PMID: 27230973 Free PMC article. Review.
-
OCT4 interprets and enhances nucleosome flexibility.Nucleic Acids Res. 2022 Oct 14;50(18):10311-10327. doi: 10.1093/nar/gkac755. Nucleic Acids Res. 2022. PMID: 36130732 Free PMC article.
-
Metastatic Phosphatase PRL-3 Induces Ovarian Cancer Stem Cell Sub-population through Phosphatase-Independent Deacetylation Modulations.iScience. 2020 Jan 24;23(1):100766. doi: 10.1016/j.isci.2019.100766. Epub 2019 Dec 12. iScience. 2020. PMID: 31887658 Free PMC article.
-
Sox2 Is an Oncogenic Driver of Small-Cell Lung Cancer and Promotes the Classic Neuroendocrine Subtype.Mol Cancer Res. 2021 Dec;19(12):2015-2025. doi: 10.1158/1541-7786.MCR-20-1006. Epub 2021 Sep 30. Mol Cancer Res. 2021. PMID: 34593608 Free PMC article.
-
Key features of the POU transcription factor Oct4 from an evolutionary perspective.Cell Mol Life Sci. 2021 Dec;78(23):7339-7353. doi: 10.1007/s00018-021-03975-8. Epub 2021 Oct 26. Cell Mol Life Sci. 2021. PMID: 34698883 Free PMC article. Review.
References
-
- Dooley T.P., Miranda,M., Jones,N.C. and DePamphilis,M.L. (1989) Transactivation of the adenovirus EIIa promoter in the absence of adenovirus E1A protein is restricted to mouse oocytes and preimplantation embryos. Development, 107, 945–956. - PubMed
-
- Gilbert S.F., (1991) Developmental Biology, 3rd Edn. Sinauer Associates, Sunderland, MA.
-
- La Thangue N.B. and Rigby,P.W.J. (1987) An adenovirus E1A-like transcription factor is regulated during the differentiation of murine embryonal carcinoma stem cells. Cell, 49, 507–513. - PubMed
-
- Kaneko K.J., and DePamphilis,M.L. (1998) Regulation of gene expression at the beginning of mammalian development and the TEAD family of transcription factors. Dev. Genet., 22, 43–55. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
