

Severe block in processing of proinsulin to insulin accompanied by elevation of des-64,65 proinsulin intermediates in islets of mice lacking prohormone convertase 1/3
- PMID: 12136131
- PMCID: PMC124908
- DOI: 10.1073/pnas.162352799
Severe block in processing of proinsulin to insulin accompanied by elevation of des-64,65 proinsulin intermediates in islets of mice lacking prohormone convertase 1/3
Abstract
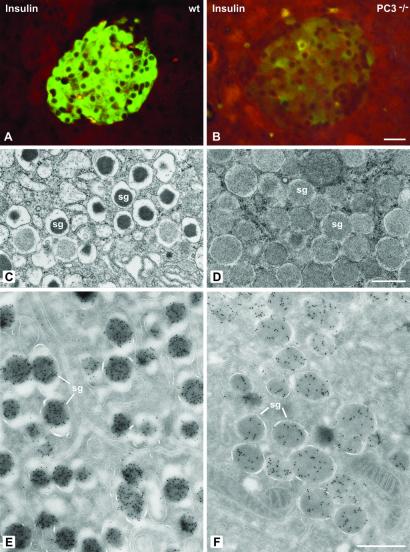
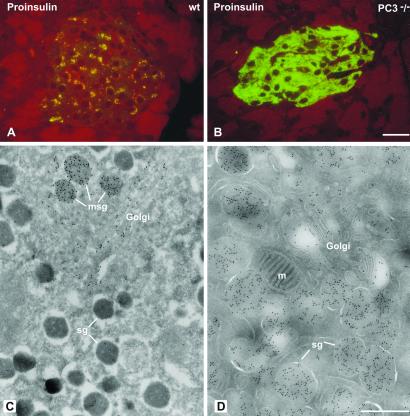
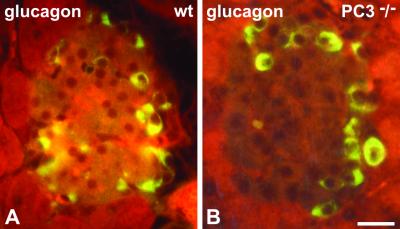
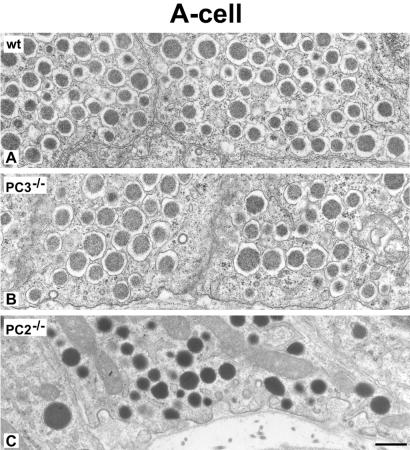
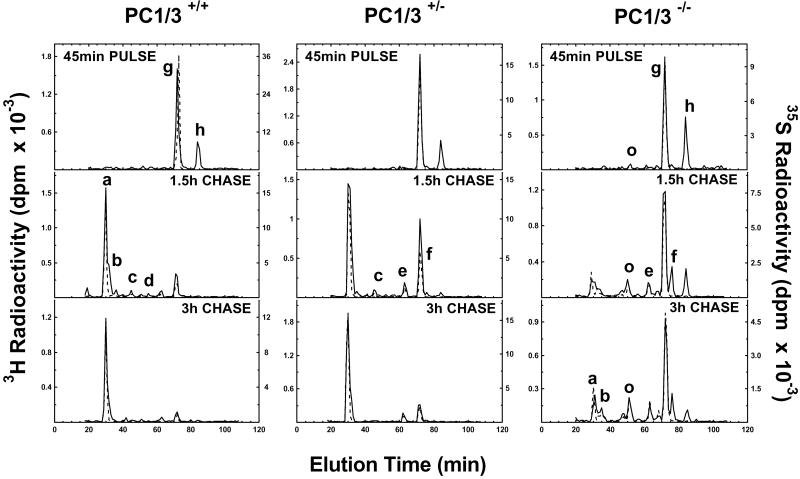
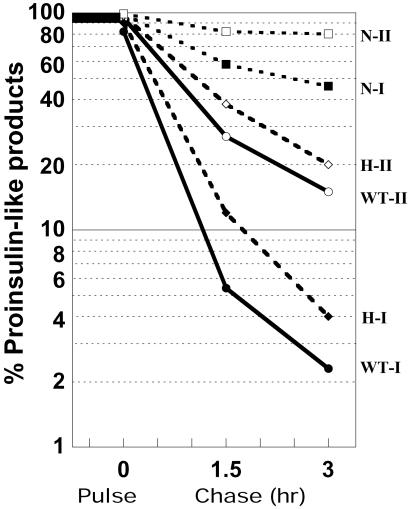
The neuroendocrine processing endoproteases PC2 and PC1/3 are expressed in the beta cells of the islets of Langerhans and participate in the processing of proinsulin to insulin and C-peptide. We have previously shown that disruption of PC2 (SPC2) expression significantly impairs proinsulin processing. Here we report that disruption of the expression of PC1/3 (SPC3) produces a much more severe block in proinsulin conversion. In nulls, pancreatic and circulating proinsulin-like components comprise 87% and 91%, respectively, of total insulin-related immunoreactivity. Heterozygotes also show a more than 2-fold elevation in proinsulin levels to approximately 12%. Immunocytochemical and ultrastructural studies of the beta cells reveal the nearly complete absence of mature insulin immunoreactivity and its replacement by that of proinsulin in abundant immature-appearing secretory granules. In contrast, alpha cell morphology and glucagon processing are normal, and there is also no defect in somatostatin-14 generation. Pulse-chase labeling studies confirm the existence of a major block in proinsulin processing in PC1/3 nulls with prolongation of half-times of conversion by 7- and 10-fold for proinsulins I and II, respectively. Lack of PC1/3 also results in increased levels of des-64,65 proinsulin intermediates generated by PC2, in contrast to PC2 nulls, in which des- 31,32 proinsulin intermediates predominate. These results confirm that PC1/3 plays a major role in processing proinsulin, but that its coordinated action with PC2 is necessary for the most efficient and complete processing of this prohormone.
Figures

References
-
- Steiner D., Chan, S. & Rubenstein, A. (2000) in Handbook of Physiology, The Endocrine System, eds. Jefferson, L. & Cherrington, A. (Oxford Univ. Press, New York), Vol. II, pp. 49–77.
-
- Steiner D. (2001) in The Enzymes, eds. Dalbey, R. E. & Sigman, D. S. (Academic, New York), Vol. XXII, pp. 163–198.
-
- Goodge K. A. & Hutton, J. C. (2000) Semin. Cell. Dev. Biol. 11, 235-242. - PubMed
-
- Zhou A., Webb, G., Zhu, X. & Steiner, D. F. (1999) J. Biol. Chem. 274, 20745-20748. - PubMed
-
- Seidah N. G. & Chrétien, M. (1999) Brain Res. 848, 45-62. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
