

Differential presentation of a soluble exogenous tumor antigen, NY-ESO-1, by distinct human dendritic cell populations
- PMID: 12138174
- PMCID: PMC124995
- DOI: 10.1073/pnas.112331099
Differential presentation of a soluble exogenous tumor antigen, NY-ESO-1, by distinct human dendritic cell populations
Abstract
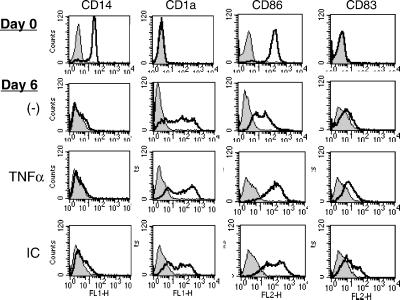
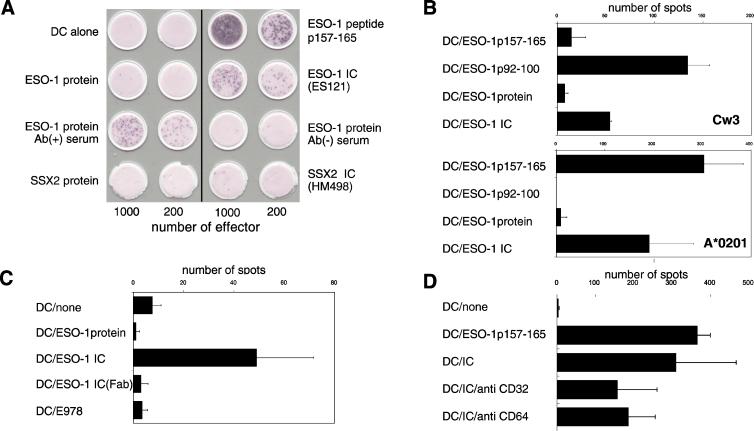
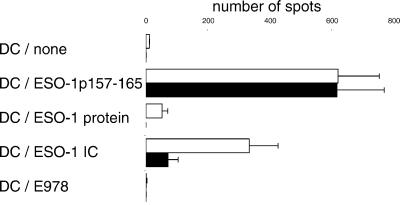
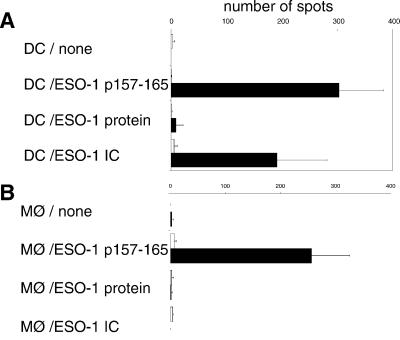
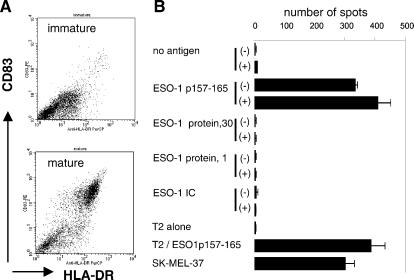
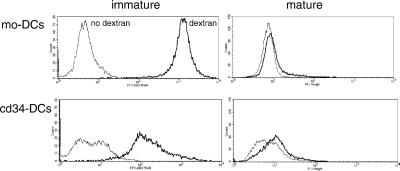
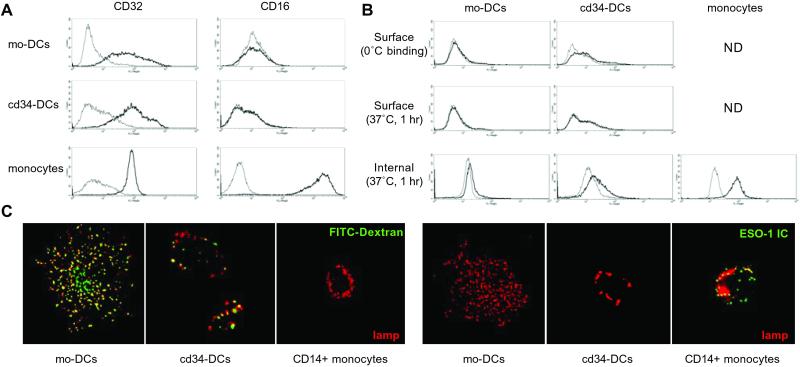
Dendritic cells (DCs) play a critical role in initiating antigen-specific immune responses, because they are able to capture exogenous antigens for presentation to naive T cells on both MHC class I and II molecules. As such, DCs represent important elements in the development of vaccine therapy for cancer. Although DCs are known to present antigens from phagocytosed tumor cells or preprocessed peptides, we explored whether they might also present soluble recombinant NY-ESO-1, a well characterized cancer antigen. We compared the abilities of human monocyte-derived DCs and DCs derived in vitro from CD34-positive stem cells to present NY-ESO-1 epitopes to MHC class I-restricted cytotoxic T cells. Although monocyte-derived DCs did not efficiently crosspresent free NY-ESO-1 protein, IgG-immune complexes containing NY-ESO-1 were avidly presented after uptake by Fcgamma receptors (FcgammaRII). In contrast, CD34-derived DCs were unable to process either soluble or immune complexed NY-ESO-1, although they efficiently presented preprocessed NY-ESO-1 peptides. This difference did not necessarily correlate with endocytic capacity. Although monocyte-derived DCs exhibited greater fluid-phase uptake than CD34-derived DCs, the two populations did not differ with respect to their surprisingly limited capacity for Fcgamma receptor-mediated endocytosis. These results indicate that monocyte-derived DCs will be easier to load by using protein antigen in vitro than CD34-derived DCs, and that the latter population exhibits a restricted ability to crosspresent soluble exogenous antigens.
Figures
Similar articles
-
Processing and cross-presentation of individual HLA-A, -B, or -C epitopes from NY-ESO-1 or an HLA-A epitope for Melan-A differ according to the mode of antigen delivery.Blood. 2010 Jul 15;116(2):218-25. doi: 10.1182/blood-2009-10-249458. Epub 2010 Apr 29. Blood. 2010. PMID: 20430956
-
Tumor antigen processing and presentation depend critically on dendritic cell type and the mode of antigen delivery.Blood. 2005 Mar 15;105(6):2465-72. doi: 10.1182/blood-2004-08-3105. Epub 2004 Nov 16. Blood. 2005. PMID: 15546948
-
Identification of a naturally processed NY-ESO-1 peptide recognized by CD8+ T cells in the context of HLA-B51.Cancer Immun. 2002 Sep 19;2:12. Cancer Immun. 2002. PMID: 12747757
-
[Development of effective antigen delivery carrier to dendritic cells via Fc receptor in cancer immunotherapy].Yakugaku Zasshi. 2007 Feb;127(2):301-6. doi: 10.1248/yakushi.127.301. Yakugaku Zasshi. 2007. PMID: 17268150 Review. Japanese.
-
Antigen capture, processing, and presentation by dendritic cells: recent cell biological studies.Hum Immunol. 1999 Jul;60(7):562-7. doi: 10.1016/s0198-8859(99)00030-0. Hum Immunol. 1999. PMID: 10426272 Review.
Cited by
-
Combined Vaccination with NY-ESO-1 Protein, Poly-ICLC, and Montanide Improves Humoral and Cellular Immune Responses in Patients with High-Risk Melanoma.Cancer Immunol Res. 2020 Jan;8(1):70-80. doi: 10.1158/2326-6066.CIR-19-0545. Epub 2019 Nov 7. Cancer Immunol Res. 2020. PMID: 31699709 Free PMC article. Clinical Trial.
-
Origin and Role of a Subset of Tumor-Associated Neutrophils with Antigen-Presenting Cell Features in Early-Stage Human Lung Cancer.Cancer Cell. 2016 Jul 11;30(1):120-135. doi: 10.1016/j.ccell.2016.06.001. Epub 2016 Jun 30. Cancer Cell. 2016. PMID: 27374224 Free PMC article.
-
Selective blockade of inhibitory Fcgamma receptor enables human dendritic cell maturation with IL-12p70 production and immunity to antibody-coated tumor cells.Proc Natl Acad Sci U S A. 2005 Feb 22;102(8):2910-5. doi: 10.1073/pnas.0500014102. Epub 2005 Feb 9. Proc Natl Acad Sci U S A. 2005. PMID: 15703291 Free PMC article.
-
Identifying baseline immune-related biomarkers to predict clinical outcome of immunotherapy.J Immunother Cancer. 2017 May 16;5:44. doi: 10.1186/s40425-017-0243-4. eCollection 2017. J Immunother Cancer. 2017. PMID: 28515944 Free PMC article. Review.
-
Prolonged antigen survival and cytosolic export in cross-presenting human gammadelta T cells.Proc Natl Acad Sci U S A. 2010 May 11;107(19):8730-5. doi: 10.1073/pnas.1002769107. Epub 2010 Apr 22. Proc Natl Acad Sci U S A. 2010. PMID: 20413723 Free PMC article.
References
-
- Banchereau J. & Steinman, R. M. (1998) Nature (London) 392, 245-252. - PubMed
-
- Mellman I. & Steinman, R. M. (2001) Cell 106, 255-258. - PubMed
-
- Albert M. L., Sauter, B. & Bhardwaj, N. (1998) Nature (London) 392, 86-89. - PubMed
-
- Norbury C. C., Hewlett, L. J., Prescott, A. R., Shastri, N. & Watts, C. (1995) Immunity 3, 783-791. - PubMed
-
- Kovacsovics-Bankowski M. & Rock, K. L. (1995) Science 267, 243-246. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
