

Steady-state nuclear localization of exportin-t involves RanGTP binding and two distinct nuclear pore complex interaction domains
- PMID: 12138183
- PMCID: PMC133969
- DOI: 10.1128/MCB.22.16.5708-5720.2002
Steady-state nuclear localization of exportin-t involves RanGTP binding and two distinct nuclear pore complex interaction domains
Abstract
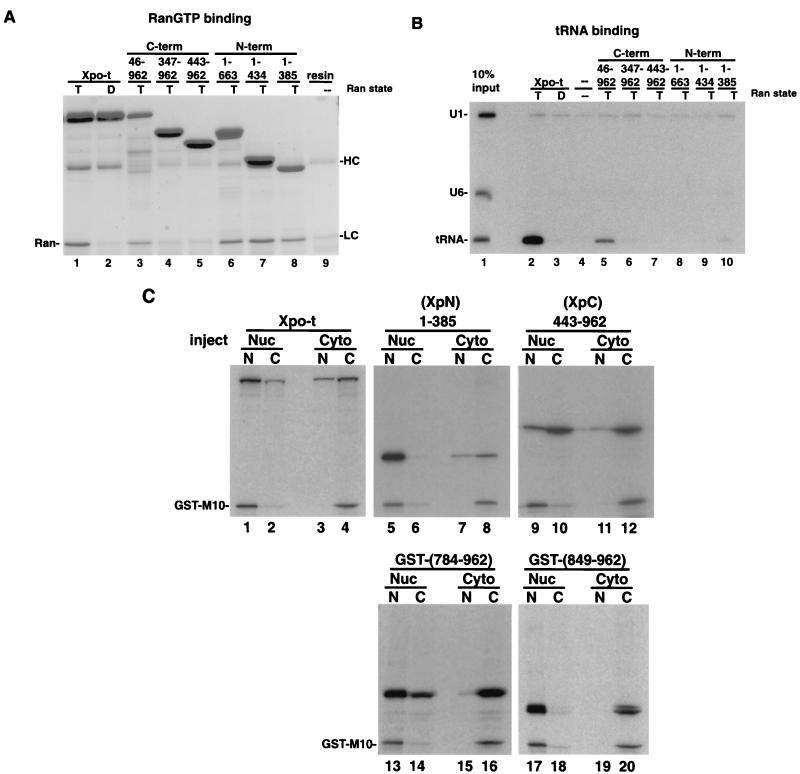
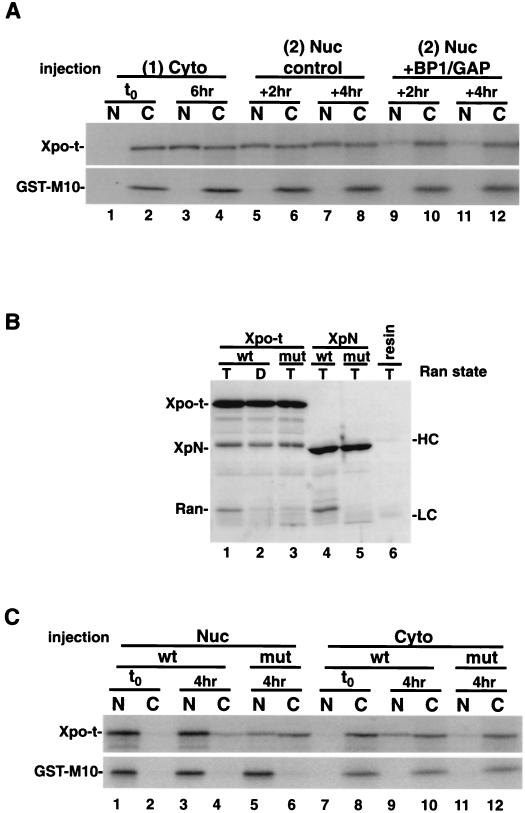
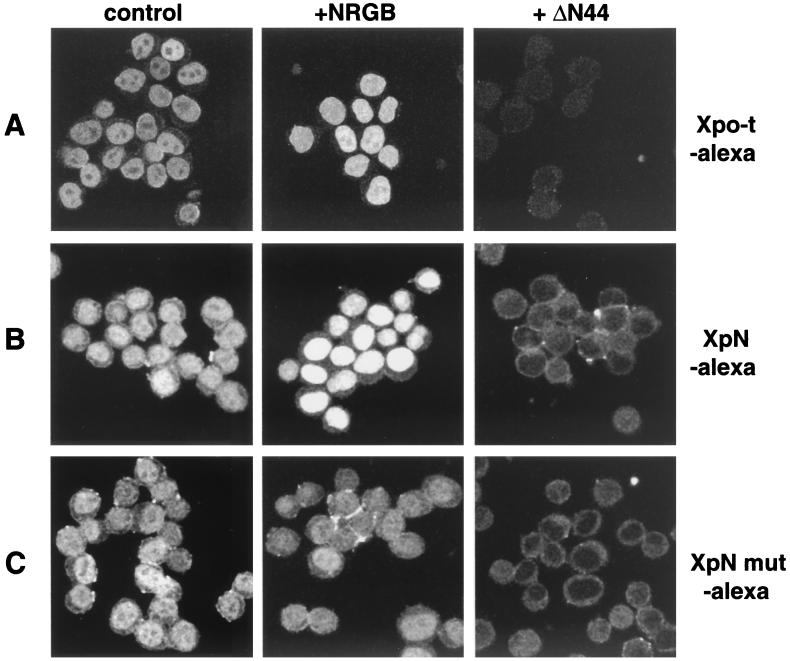
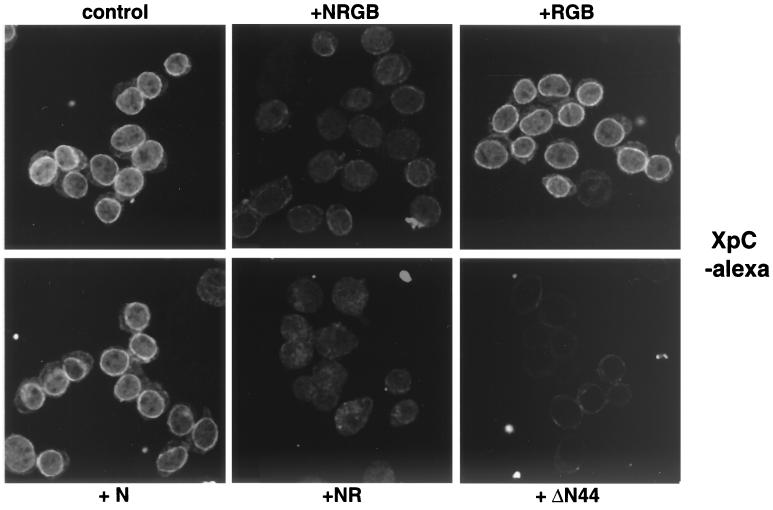
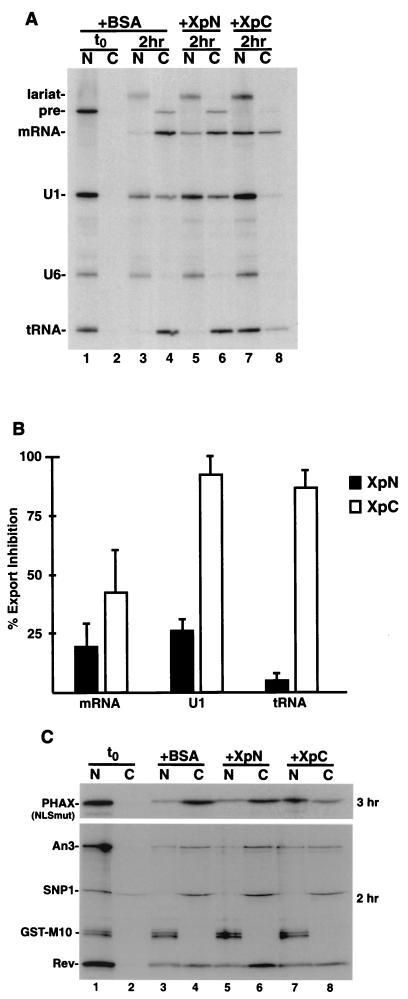
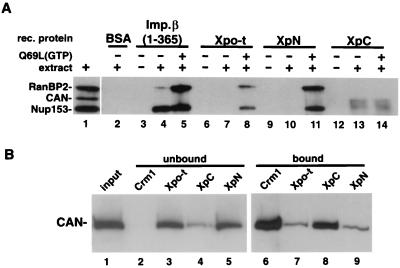
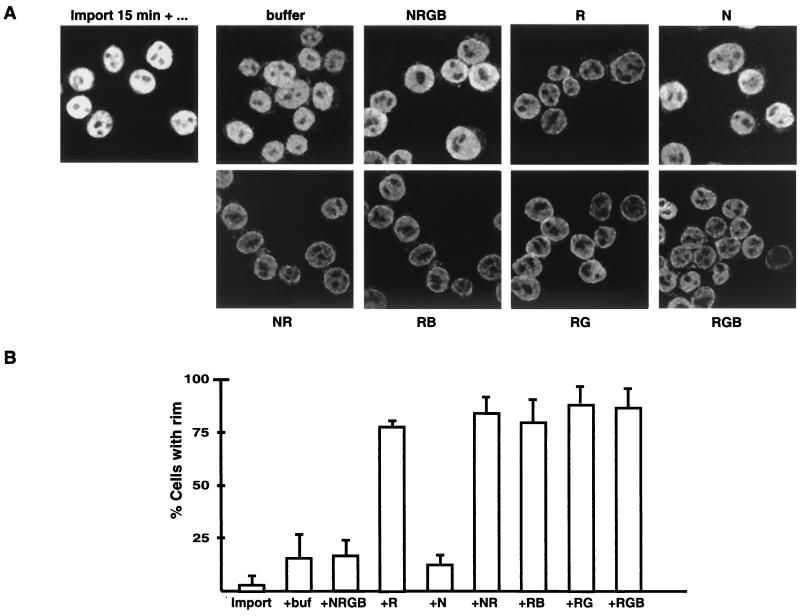
Vertebrate tRNA export receptor exportin-t (Xpo-t) binds to RanGTP and mature tRNAs cooperatively to form a nuclear export complex. Xpo-t shuttles bidirectionally through nuclear pore complexes (NPCs) but is mainly nuclear at steady state. The steady-state distribution of Xpo-t is shown to depend on its interaction with RanGTP. Two distinct Xpo-t NPC interaction domains that bind differentially to peripherally localized nucleoporins in vitro are identified. The N terminus binds to both Nup153 and RanBP2/Nup358 in a RanGTP-dependent manner, while the C terminus binds to CAN/Nup214 independently of Ran. We propose that these interactions increase the concentration of tRNA export complexes and of empty Xpo-t in the vicinity of NPCs and thus increase the efficiency of the Xpo-t transport cycle.
Figures


Similar articles
-
The role of exportin-t in selective nuclear export of mature tRNAs.EMBO J. 1998 Dec 15;17(24):7430-41. doi: 10.1093/emboj/17.24.7430. EMBO J. 1998. PMID: 9857198 Free PMC article.
-
Identification of a tRNA-specific nuclear export receptor.Mol Cell. 1998 Feb;1(3):359-69. doi: 10.1016/s1097-2765(00)80036-2. Mol Cell. 1998. PMID: 9660920
-
Recognition of engineered tRNAs with an extended 3' end by Exportin-t (Xpo-t) and transport of tRNA-attached ribozymes to the cytoplasm in somatic cells.Biomacromolecules. 2001 Winter;2(4):1229-42. doi: 10.1021/bm0101062. Biomacromolecules. 2001. PMID: 11777397
-
Nucleocytoplasmic transport enters the atomic age.Curr Opin Cell Biol. 2001 Jun;13(3):310-9. doi: 10.1016/s0955-0674(00)00213-1. Curr Opin Cell Biol. 2001. PMID: 11343901 Review.
-
Mechanistic Insights from Structural Analyses of Ran-GTPase-Driven Nuclear Export of Proteins and RNAs.J Mol Biol. 2016 May 22;428(10 Pt A):2025-39. doi: 10.1016/j.jmb.2015.09.025. Epub 2015 Oct 28. J Mol Biol. 2016. PMID: 26519791 Review.
Cited by
-
Altered nuclear tRNA metabolism in La-deleted Schizosaccharomyces pombe is accompanied by a nutritional stress response involving Atf1p and Pcr1p that is suppressible by Xpo-t/Los1p.Mol Biol Cell. 2012 Feb;23(3):480-91. doi: 10.1091/mbc.E11-08-0732. Epub 2011 Dec 7. Mol Biol Cell. 2012. PMID: 22160596 Free PMC article.
-
Structures of the tRNA export factor in the nuclear and cytosolic states.Nature. 2009 Sep 3;461(7260):60-5. doi: 10.1038/nature08394. Nature. 2009. PMID: 19680239
-
The Ran Pathway in Drosophila melanogaster Mitosis.Front Cell Dev Biol. 2015 Nov 26;3:74. doi: 10.3389/fcell.2015.00074. eCollection 2015. Front Cell Dev Biol. 2015. PMID: 26636083 Free PMC article. Review.
-
Distinct functional domains within nucleoporins Nup153 and Nup98 mediate transcription-dependent mobility.Mol Biol Cell. 2004 Apr;15(4):1991-2002. doi: 10.1091/mbc.e03-10-0743. Epub 2004 Jan 12. Mol Biol Cell. 2004. PMID: 14718558 Free PMC article.
-
Exportins can inhibit major mitotic assembly events in vitro: membrane fusion, nuclear pore formation, and spindle assembly.Nucleus. 2020 Dec;11(1):178-193. doi: 10.1080/19491034.2020.1798093. Nucleus. 2020. PMID: 32762441 Free PMC article.
References
-
- Allen, N. P. C., L. Huang, A. Burlingame, and M. Rexach. 2001. Proteomic analysis of nucleoporin interacting proteins. J. Biol. Chem. 276:29268-29274. - PubMed
-
- Arts, G.-J., M. Fornerod, and I. W. Mattaj. 1998. Identification of a nuclear export receptor for tRNA. Curr. Biol. 8:305-314. - PubMed
-
- Bayliss, R., T. Littlewood, and M. Stewart. 2000. Structural basis for the interaction between FxFG nucleoporin repeats and importin-beta in nuclear trafficking. Cell 102:99-108. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous