

Cluster II che genes from Pseudomonas aeruginosa are required for an optimal chemotactic response
- PMID: 12142407
- PMCID: PMC135244
- DOI: 10.1128/JB.184.16.4374-4383.2002
Cluster II che genes from Pseudomonas aeruginosa are required for an optimal chemotactic response
Abstract
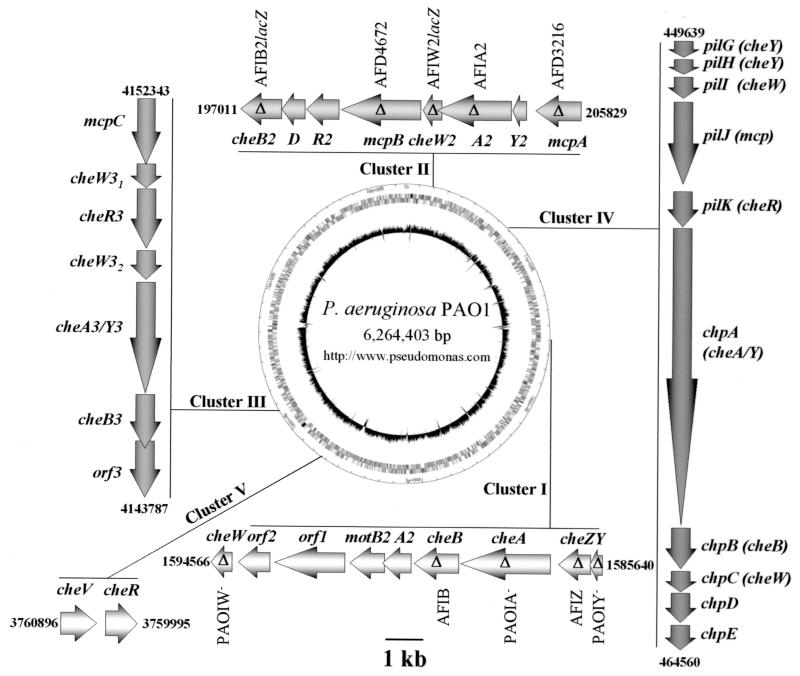
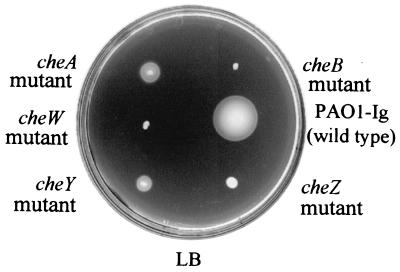
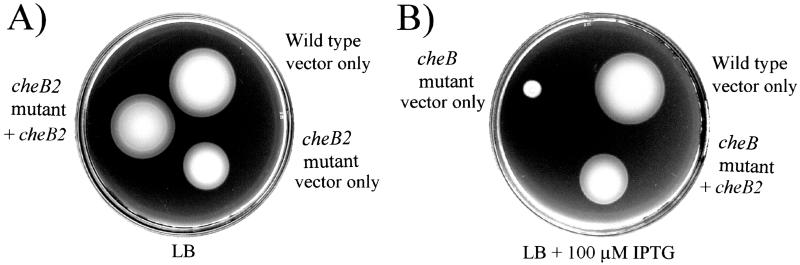
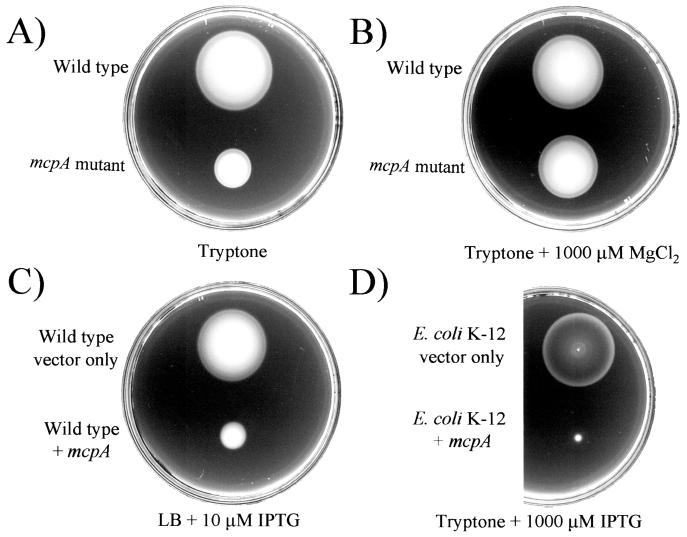
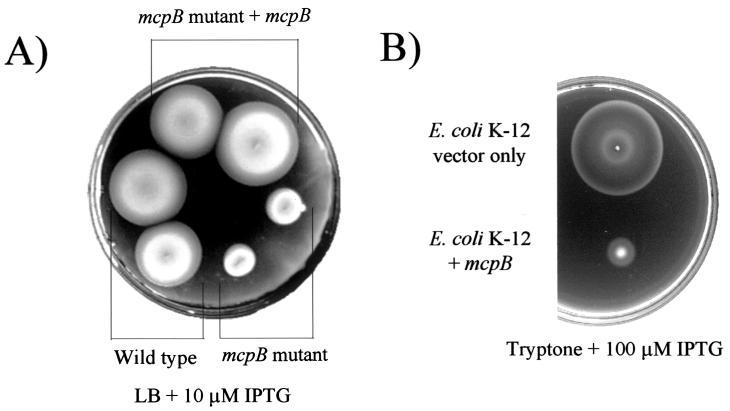
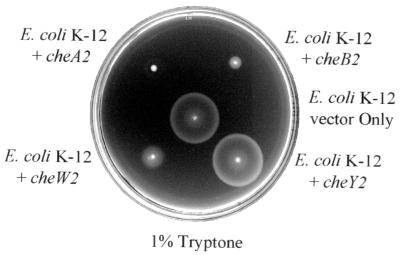
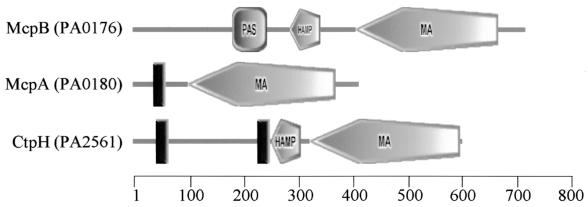
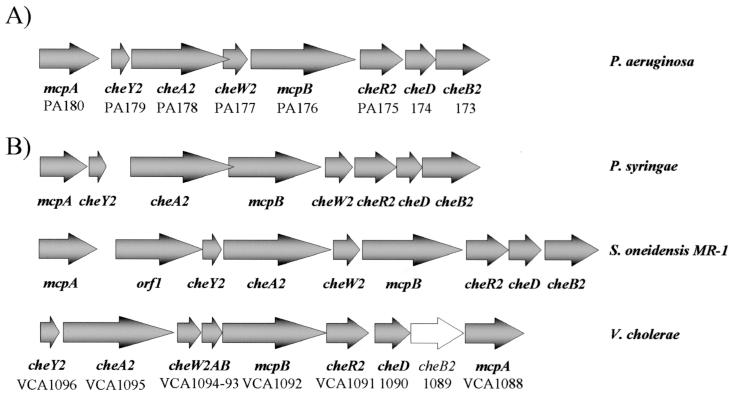
Pseudomonas aeruginosa, a gamma-proteobacterium, is motile by means of a single polar flagellum and is chemotactic to a variety of organic compounds and phosphate. P. aeruginosa has multiple homologues of Escherichia coli chemotaxis genes that are organized into five gene clusters. Previously, it was demonstrated that genes in cluster I and cluster V are essential for chemotaxis. A third cluster (cluster II) contains a complete set of che genes, as well as two genes, mcpA and mcpB, encoding methyl-accepting chemotaxis proteins. Mutations were constructed in several of the cluster II che genes and in the mcp genes to examine their possible contributions to P. aeruginosa chemotaxis. A cheB2 mutant was partially impaired in chemotaxis in soft-agar swarm plate assays. Providing cheB2 in trans complemented this defect. Further, overexpression of CheB2 restored chemotaxis to a completely nonchemotactic, cluster I, cheB-deficient strain to near wild-type levels. An mcpA mutant was defective in chemotaxis in media that were low in magnesium. The defect could be relieved by the addition of magnesium to the swarm plate medium. An mcpB mutant was defective in chemotaxis when assayed in dilute rich soft-agar swarm medium or in minimal-medium swarm plates containing any 1 of 60 chemoattractants. The mutant phenotype could be complemented by the addition of mcpB in trans. Overexpression of either McpA or McpB in P. aeruginosa or Escherichia coli resulted in impairment of chemotaxis, and these cells had smooth-swimming phenotypes when observed under the microscope. Expression of P. aeruginosa cheA2, cheB2, or cheW2 in E. coli K-12 completely disrupted wild-type chemotaxis, while expression of cheY2 had no effect. These results indicate that che cluster II genes are expressed in P. aeruginosa and are required for an optimal chemotactic response.
Figures
References
-
- Aravind, L., and C. P. Ponting. 1999. The cytoplasmic helical linker domain of receptor histidine kinase and methyl-accepting proteins is common to many prokaryotic signaling proteins. FEMS Microbiol. Lett. 176:111-116. - PubMed
-
- Armitage, J. P., and R. Schmitt. 1997. Bacterial chemotaxis: Rhodobacter sphaeroides and Sinorhizobium meliloti—variations on a theme? Microbiology 143:3671-3682. - PubMed
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, K. Struhl, L. M. Albright, D. M. Coen, and A. Varki (ed.). 1998. Current protocols in molecular biology, vol. 1. John Wiley and Sons Inc., New York, N.Y.
-
- Bachmann, B. J. 1987. Linkage map of Escherichia coli K-12, 7th ed., p. 807-876. In F. C. Neidhardt, J. L. Ingraham, K. B. Low, B. Magasanik, M. Schaechter, and H. E. Umbarger (ed.), Escherichia coli and Salmonella typhimurium: cellular and molecular biology, vol. 2. American Society for Microbiology, Washington, D.C.
-
- Bourret, R. B., and A. M. Stock. 2002. Molecular information processing: lessons from bacterial chemotaxis. J. Biol. Chem. 277:9625-9628. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
