

icaR encodes a transcriptional repressor involved in environmental regulation of ica operon expression and biofilm formation in Staphylococcus epidermidis
- PMID: 12142410
- PMCID: PMC135245
- DOI: 10.1128/JB.184.16.4400-4408.2002
icaR encodes a transcriptional repressor involved in environmental regulation of ica operon expression and biofilm formation in Staphylococcus epidermidis
Abstract
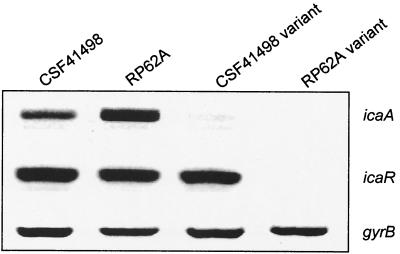
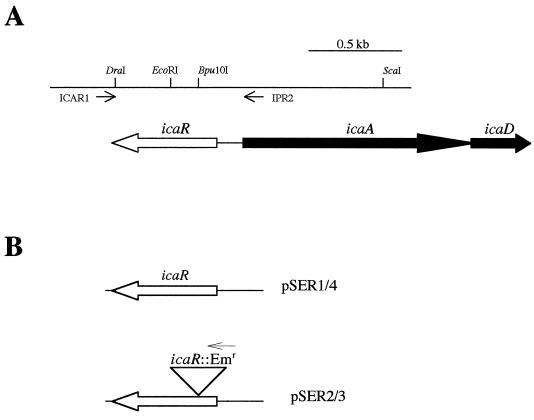
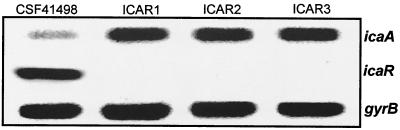
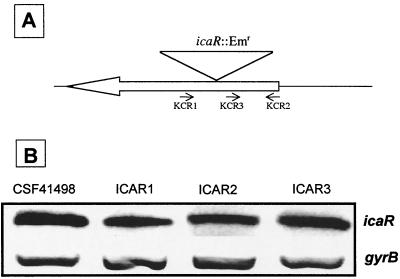
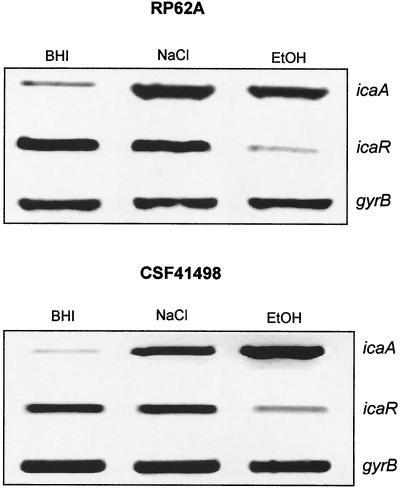
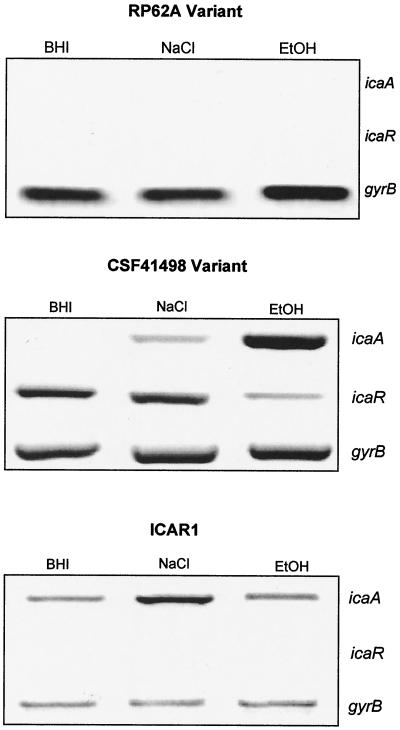
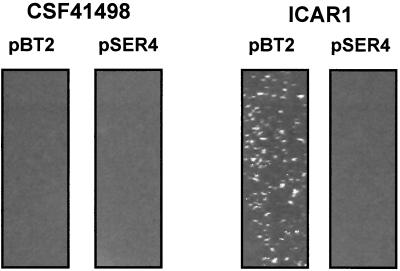
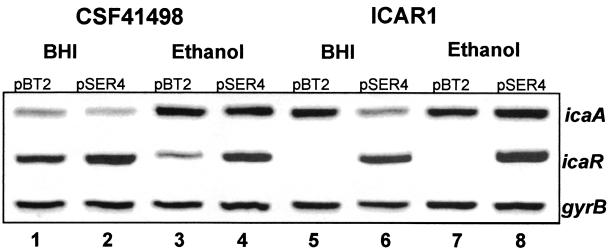
Biofilm formation in Staphylococcus epidermidis is dependent upon the ica operon-encoded polysaccharide intercellular adhesin, which is subject to phase-variable and environmental regulation. The icaR gene, located adjacent to the ica operon, appears to be a member of the tetR family of transcriptional regulators. In the reference strain RP62A, reversible inactivation of the ica operon by IS256 accounts for 25 to 33% of phase variants. In this study, icaA and icaR regulation were compared in RP62A and a biofilm-forming clinical isolate, CSF41498, in which IS256 is absent. Predictably, ica operon expression was detected only in wild-type CSF41498 and RP62A but not in non-IS256-generated phase variants. In contrast, the icaR gene was not expressed in RP62A phase variants but was expressed in CSF41498 variants. An icaR::Em(r) insertion mutation in CSF41498 resulted in an at least a 5.8-fold increase in ica operon expression but did not significantly alter regulation of the icaR gene itself. Activation of ica operon transcription by ethanol in CSF41498 was icaR dependent. In contrast, a small but significant induction of ica by NaCl and glucose (NaCl-glucose) was observed in the icaR::Em(r) mutant. In addition, transcription of the icaR gene itself was not significantly affected by NaCl-glucose but was repressed by ethanol. Expression of the ica operon was induced by ethanol or NaCl-glucose in phase variants of CSF41498 (icaR+) but not in RP62A variants (icaR deficient). These data indicate that icaR encodes a repressor of ica operon transcription required for ethanol but not NaCl-glucose activation of ica operon expression and biofilm formation.
Figures
References
-
- Aramaki, H., N. Yagi, and M. Suzuki. 1995. Residues important for the function of a multihelical DNA binding domain in the new transcription factor family of Cam and Tet repressors. Protein Eng. 8:1259-1266. - PubMed
-
- Arciola, C. R., S. Collamati, E. Donati, and L. Montanaro. 2001. A rapid PCR method for the detection of slime-producing strains of Staphylococcus epidermidis and S. aureus in periprosthesis infections. Diagn. Mol. Pathol. 10:130-137. - PubMed
-
- Bruckner, R. 1997. Gene replacement in Staphylococcus carnosus and Staphylococcus xylosus. FEMS Microbiol. Lett. 151:1-8. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
