

Thermal denaturation of Bungarus fasciatus acetylcholinesterase: Is aggregation a driving force in protein unfolding?
- PMID: 12142456
- PMCID: PMC2373691
- DOI: 10.1110/ps.0205102
Thermal denaturation of Bungarus fasciatus acetylcholinesterase: Is aggregation a driving force in protein unfolding?
Abstract
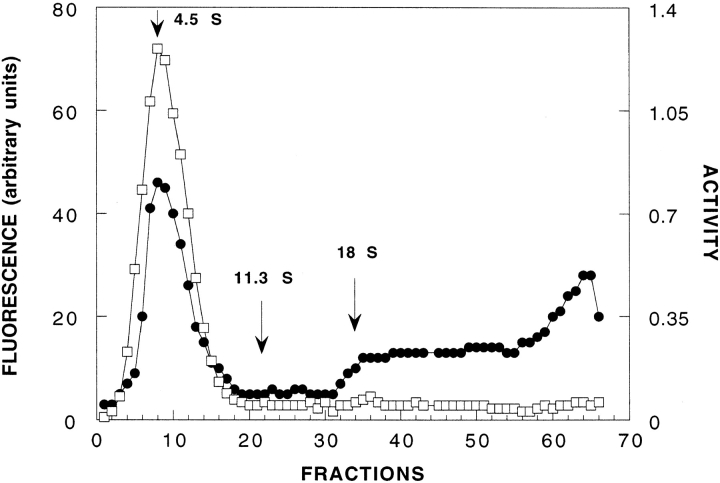
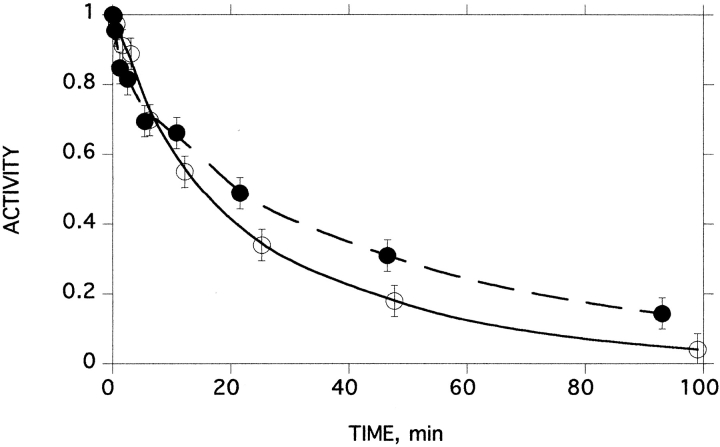
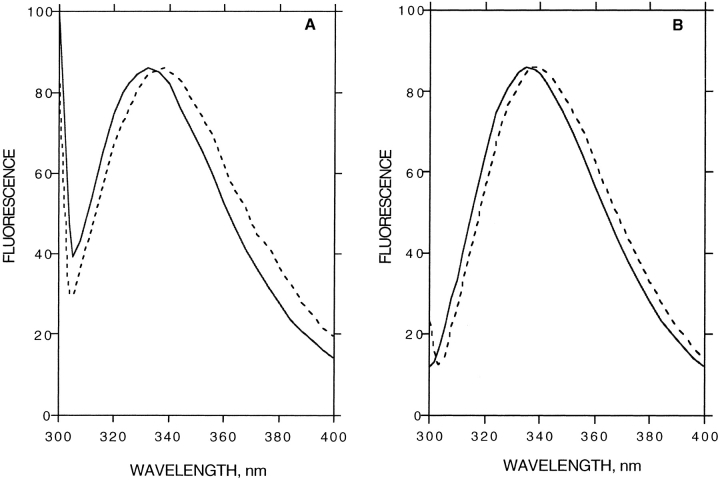
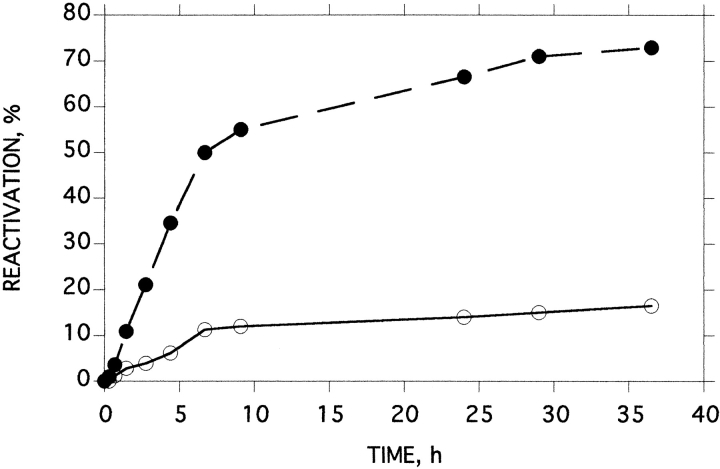
A monomeric form of acetylcholinesterase from the venom of Bungarus fasciatus is converted to a partially unfolded molten globule species by thermal inactivation, and subsequently aggregates rapidly. To separate the kinetics of unfolding from those of aggregation, single molecules of the monomeric enzyme were encapsulated in reverse micelles of Brij 30 in 2,2,4-trimethylpentane, or in large unilamellar vesicles of egg lecithin/cholesterol at various protein/micelle (vesicle) ratios. The first-order rate constant for thermal inactivation at 45 degrees C, of single molecules entrapped within the reverse micelles (0.031 min(-1)), was higher than in aqueous solution (0.007 min(-1)) or in the presence of normal micelles (0.020 min(-1)). This clearly shows that aggregation does not provide the driving force for thermal inactivation of BfAChE. Within the large unilamellar vesicles, at average protein/vesicle ratios of 1:1 and 10:1, the first-order rate constants for thermal inactivation of the encapsulated monomeric acetylcholinesterase, at 53 degrees C, were 0.317 and 0.342 min(-1), respectively. A crosslinking technique, utilizing the photosensitive probe, hypericin, showed that thermal denaturation produces a distribution of species ranging from dimers through to large aggregates. Consequently, at a protein/vesicle ratio of 10:1, aggregation can occur upon thermal denaturation. Thus, these experiments also demonstrate that aggregation does not drive the thermal unfolding of Bungarus fasciatus acetylcholinesterase. Our experimental approach also permitted monitoring of recovery of enzymic activity after thermal denaturation in the absence of a competing aggregation process. Whereas no detectable recovery of enzymic activity could be observed in aqueous solution, up to 23% activity could be obtained for enzyme sequestered in the reverse micelles.
Figures








References
-
- Berne, B. and Pecora, R. 1976. Dynamic light scattering. Wiley, New York.
-
- Bhattacharyya, K. and Basak, S. 1993. Fluorescence study of melanocyte stimulating hormones in AOT reverse micelles. Biophys. Chem. 47 21–31. - PubMed
-
- Bochkareva, E.S., Lissin N.M., Flynn, G.C., Rothman, J.E., and Girshovich, A.S. 1992. Positive cooperativity in the functioning of molecular chaperone GroEL. J. Biol. Chem. 267 6796–6800. - PubMed
-
- Boukobza, E., Sonnenfeld, A., and Haran, G. 2001. Immobilization in surface-tethered lipd vesicles as a new tool for single biomolecule spectroscopy. J. Phys. Chem. B 105 12165–12170.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
