

doi: 10.1126/science.1069076.
Helicobacter pylori SabA adhesin in persistent infection and chronic inflammation
Affiliations
- PMID: 12142529
- PMCID: PMC2570540
- DOI: 10.1126/science.1069076
Item in Clipboard
Helicobacter pylori SabA adhesin in persistent infection and chronic inflammation
Science.
.
Abstract
Helicobacter pylori adherence in the human gastric mucosa involves specific bacterial adhesins and cognate host receptors. Here, we identify sialyl-dimeric-Lewis x glycosphingolipid as a receptor for H. pylori and show that H. pylori infection induced formation of sialyl-Lewis x antigens in gastric epithelium in humans and in a Rhesus monkey. The corresponding sialic acid-binding adhesin (SabA) was isolated with the "retagging" method, and the underlying sabA gene (JHP662/HP0725) was identified. The ability of many H. pylori strains to adhere to sialylated glycoconjugates expressed during chronic inflammation might thus contribute to virulence and the extraordinary chronicity of H. pylori infection.
Figures
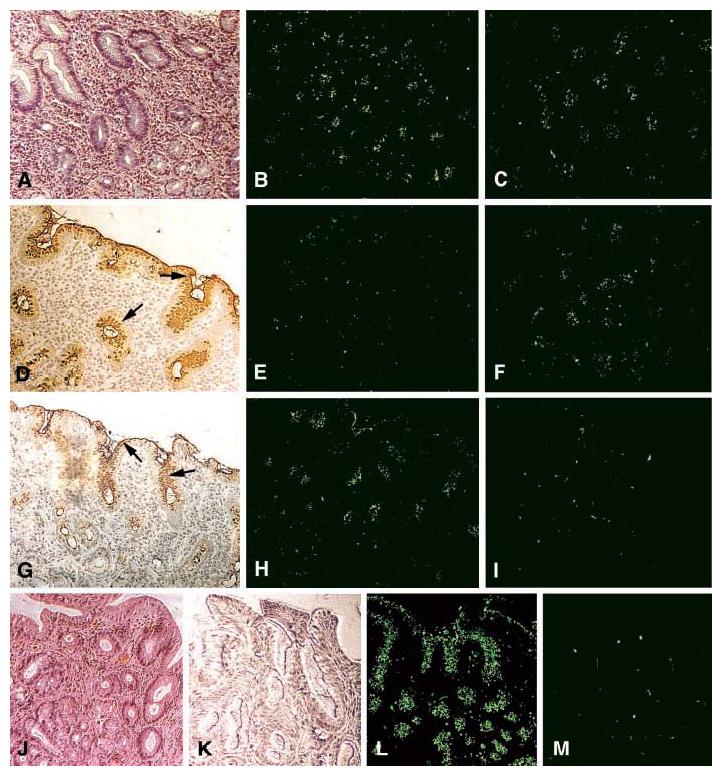
The sLex antigen confers adherence of H. pylori to the epithelium of H. pylori–infected (strain WU12) human gastric mucosa (Fig. 3A). H/E staining reveals mucosal inflammation (A). The 17875 parent strain (B) and the babA1A2 mutant (C) both adhere to the gastric epithelium. The surface epithelium stains positive (arrows) with both the Leb mAb (D) and sLex mAb (G) [AIS described in (14)]. The 17875 strain and babA1A2 mutant responded differently after pretreatment (inhibition) with soluble Leb antigen [(E) and (F), respectively)], or with soluble sLex antigen [(H) and (I), respectively)]. In conclusion, the Leb antigen blocked binding of the 17875 strain, whereas the sLex antigen blocked binding of the babA1A2 mutant. H/E-stained biopsy with no H. pylori infection (J). No staining was detected with the sLex mAb (K). Here, strain 17875 adhered (L), in contrast to the babA1A2 mutant (M), because noninflamed gastric mucosa is low in sialylation (K).
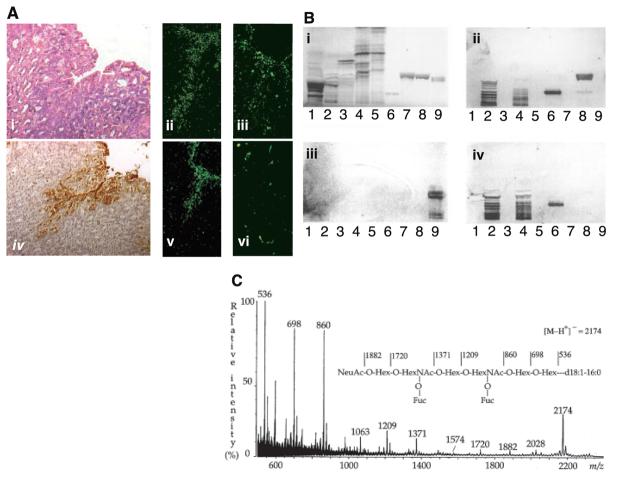
(A) The sLex antigen confers adherence of H. pylori to the transgenic Leb mouse gastric mucosa. H/E-stained biopsy of Leb mouse gastric mucosa (i). The 17875 strain (ii) and babA1A2 mutant (iii) both adhered to the surface epithelium, which stained positive with the sLex mAb (iv). Adherence by the babA1A2 mutant was lost (vi), while adherence by the 17875 strain was unperturbed (v), after pretreatment of the bacteria with soluble sLex antigen. (B) H. pylori binds to fucosylated gangliosides such as the sLex GSL. GSLs were separated on TLC and chemically stained (i) (14). Chromatograms were probed with sLex mAb (ii), the 17875 strain (iii), and the babA1A2 mutant (iv). Lanes contained acid (i.e., sialylated) GSLs of calf brain, 40 μg; acid GSLs of human neutrophil granulocytes, 40 μg; sample number 2 after desialylation, 40 μg; acid GSLs of adenocarcinoma, 40 μg; sample number 4 after desialylation, 40 μg; sdiLex GSL, 1 μg; sLea GSL, 4 μg; s(mono)Lex GSL, 4 μg; and Leb GSL, 4 μg. Binding results are summarized in table S1. (C) The high-affinity H. pylori GSL receptor was structurally identified by negative ion fast atom bombardment mass spectrometry. The molecular ion (M-H+)− at m/z 2174 indicates a GSL with one NeuAc, two fucoses, two N-acetylhexosamines, four hexoses, and d18 :1-16:0; and the sequence NeuAcHex(Fuc)HexNAc-Hex(Fuc)HexNAcHexHex was deduced. 1H-NMR spectroscopy resolved the sdiLex antigen; NeuAcα2.3Galβ1.4(Fucα1.3)GlcNAcβ1.3Galβ1.4(Fucα1.3) GlcNAcβ1.3Galβ1.4Glcβ1Cer (14).
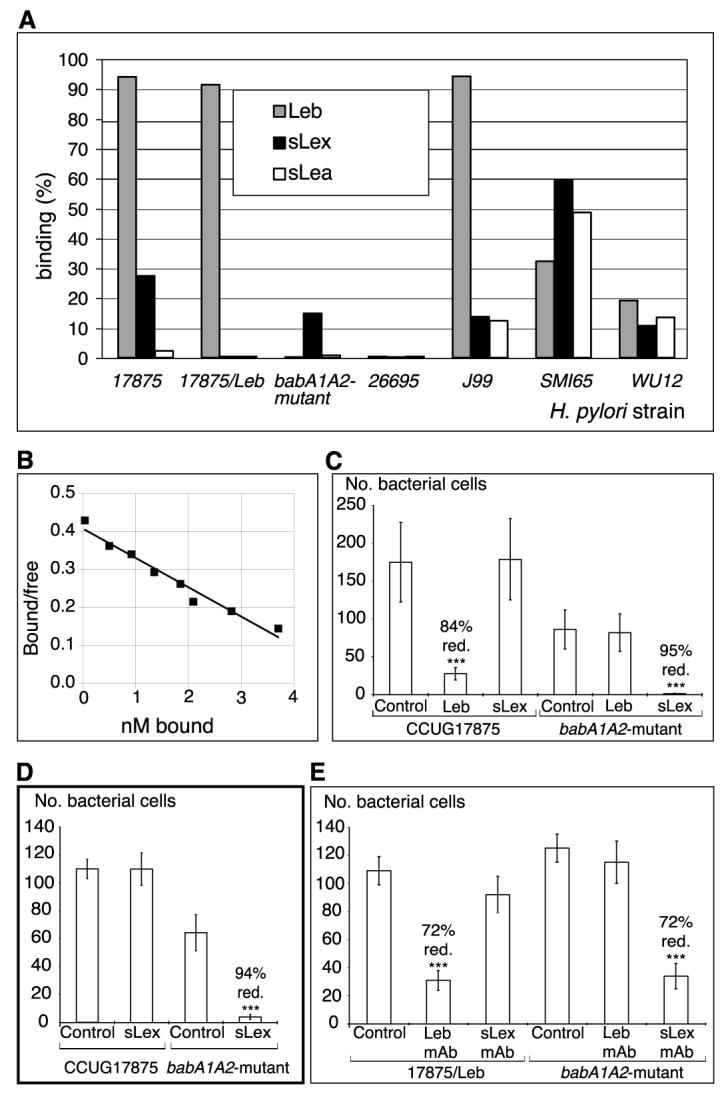
H. pylori binds sialylated antigens. (A) H. pylori strains and mutants (14) were analyzed for binding to different 125I-labeled soluble fucosylated and sialylated (Lewis) antigen-conjugates [RIA in (14)]. The bars give bacterial binding, and conjugates used are given in the diagram. (B) For affinity analyses (16), the sLex conjugate was added in titration series. The babA1A2 mutant was incubated for 3 hours with the s(mono)Lex conjugate to allow for equilibrium in binding, which demonstrates an affinity (Ka) of 1 × 108 M−1. (C) Adherence of strain 17875 (sLex and Leb-antigen binding) and babA1A2 mutant to biopsy with inflammation and H. pylori infection, as scored by the number of bound bacteria after pretreatment with soluble Leb (Fig. 1, E and F) or sLex antigen (Fig. 1, H and I) (14). The Leb antigen reduced adherence of strain 17875 by >80%, whereas the sLex antigen abolished adherence of the babA1A2 mutant. (D) Adherence of strain 17875 and babA1A2 mutant to biopsy of Leb mouse gastric mucosa was scored by the number of bound bacteria after pretreatment with sLex conjugate. Adherence by the babA1A2 mutant was abolished (Fig. 2A, vi), while adherence by strain 17875 was unperturbed (Fig. 2A, v). (E) Adherence of spontaneous phase variant strain 17875/Leb (Leb-antigen binding only in Fig. 3A) and babA1A2 mutant was analyzed after pretreatment of histo-sections of human gastric mucosa with mAbs recognizing the Leb antigen or the sdiLex antigen (FH6), with efficient inhibition of adherence of both strain 17875/Leb and the babA1A2 mutant, respectively. Value P < 0.001 (***), value P < 0.01 (**), value P < 0.05 (*).
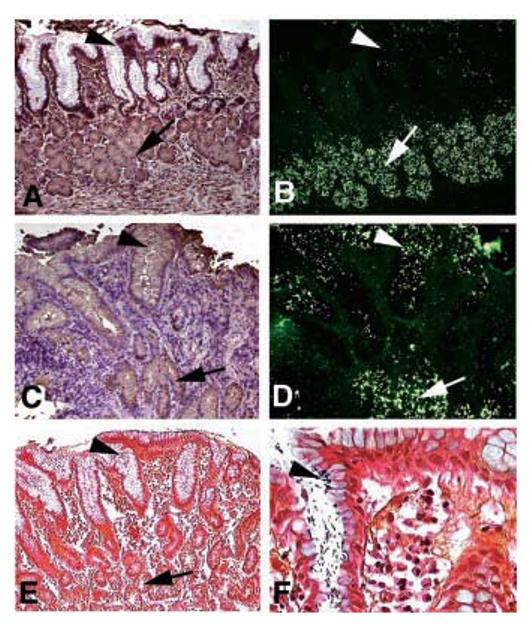
Infection induces expression of sLex antigens that serve as binding sites for H. pylori adherence. (A) and (B) came from a Rhesus monkey with healthy gastric mucosa. (C)to(F) came from the same Rhesus monkey 9 months after established reinfection with virulent clinical H. pylori isolates. Sections were analyzed with the sLex mAb (A) and (C) and by adherence in situ with the babA1A2 mutant (B) and (D). In the healthy animal, both the sLex mAb staining and adherence of the babA1A2 mutant is restricted to the deeper gastric glands [(A) and (B), arrows] [i.e., no mAb-staining of sLex antigens and no babA1A2-mutant bacteria present in the surface epithelium (A) and (B), arrowhead]. In contrast, in the H. pylori–infected and inflamed gastric mucosa, the sLex antigen is heavily expressed in the surface epithelium [(C), arrowhead with brownish immunostaining], in addition to the deeper gastric glands [(C), arrow]. The up-regulated sLex antigen now supports massive colocalized adherence of the babA1A2 mutant in situ [(D), ar- rowhead] in addition to the constant binding to the deeper gastric glands [(B) and (D), arrow]. The established infection was visualized by Genta stain, where the H. pylori microbes were present in the surface epithelium [(E) and (F), arrowhead; (F) is at a higher magnification], while no microbes were detected in the deeper gastric glands [(E), arrow].
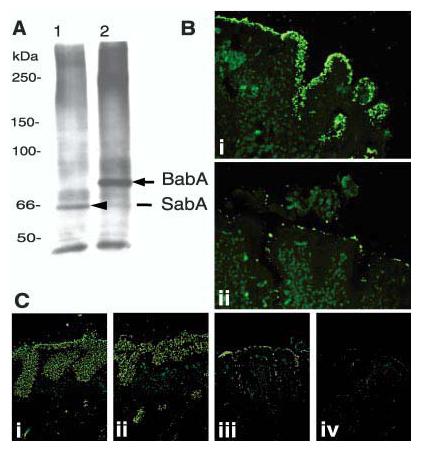
Retagging of SabA and identification of the corresponding gene, sabA. After contact-dependent retagging (of the babA1A2 mutant) with crosslinker labeled sLex conjugate, the 66-kDa biotin-tagged adhesin SabA [(A), lane 1] was identified by SDS–polyacrylamide gel electrophoresis/steptavidin-blot, magnetic-bead-purified, and analyzed by MS for peptide masses (14). As a control, the Leb conjugate was used to retag 17875 bacteria, which visualized the 75-kDa BabA adhesin [(A), lane 2] (4). (B) The J99/JHP662::cam (sabA) mutant does not bind the sLex antigen but adheres to human gastric epithelium [(B), i], due to the BabA-adhesin, because pretreatment with soluble Leb antigen inhibited binding to the epithelium [(B), ii]. (C) The J99 wild-type strain (which binds both Leb and sLex antigens) [(C), i] and the J99 sabA mutant (which binds Leb antigen only) [(C), ii] both exhibit strong adherence to the gastric mucosa. In comparison, the J99 babA mutant binds by lower-affinity interactions [(C), iii], while the J99 sabAbabA mutant, which is devoid of both adherence properties, does not bind to the gastric mucosa [(C), iv].
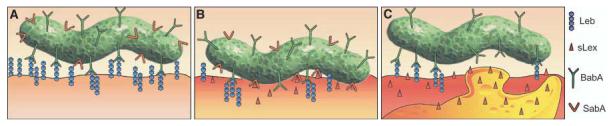
H. pylori adherence in health and disease. This figure illustrates the proficiency of H. pylori for adaptive multistep mediated attachment. (A) H. pylori (in green) adherence is mediated by the Leb blood group antigen expressed in glycoproteins (blue chains) in the gastric surface epithelium (the lower surface) (3, 32). H. pylori uses BabA (green Y's) for strong and specific recognition of the Leb antigen (4). Most of the sLex-binding isolates also bind the Leb antigen (SabA, in red Y's). (B) During persistent infection and chronic inflammation (gastritis), H. pylori triggers the host tissue to retailor the gastric mucosal glycosylation patterns to up-regulate the inflammation-associated sLex antigens (red host, triangles). Then, SabA (red Y structures) performs Selectin-mimicry by binding the sialyl-(di)-Lewis x/a glycosphingolipids, for membrane close attachment and apposition. (C) At sites of vigorous local inflammatory response, as illustrated by the recruited activated white blood cell (orange “bleb”), those H. pylori subclones that have lost sLex-binding capacity due to ON/OFF frameshift mutation might have gained local advantage in the prepared escaping of intimate contact with (sialylated) lymphocytes or other defensive cells. Such adaptation of bacterial adherence properties and subsequent inflammation pressure could be major contributors to the extraordinary chronicity of H. pylori infection in human gastric mucosa.
References
-
- Cover TL, et al. In: Principles of Bacterial Pathogenesis. Groisman EA, editor. Academic Press; New York: 2001. pp. 509–558.
-
- Gerhard M, et al. In: Helicobacter pylori: Molecular and Cellular Biology. Suerbaum S, Achtman M, editors. Horizon Scientific Press; Norfolk, UK: 2001. chap. 12.
-
- Borén T, et al. Science. 1993;262:1892. - PubMed
-
- Ilver D, et al. Science. 1998;279:373. - PubMed
-
- Hurtig M, Borén T. in preparation.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
