

Yhh1p/Cft1p directly links poly(A) site recognition and RNA polymerase II transcription termination
- PMID: 12145212
- PMCID: PMC126137
- DOI: 10.1093/emboj/cdf390
Yhh1p/Cft1p directly links poly(A) site recognition and RNA polymerase II transcription termination
Abstract
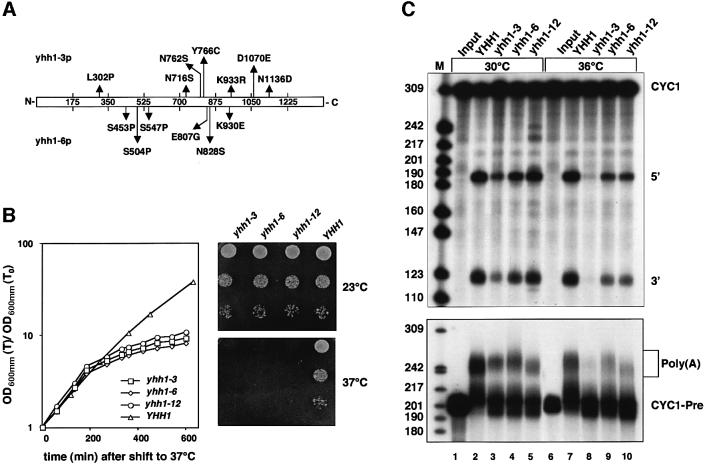
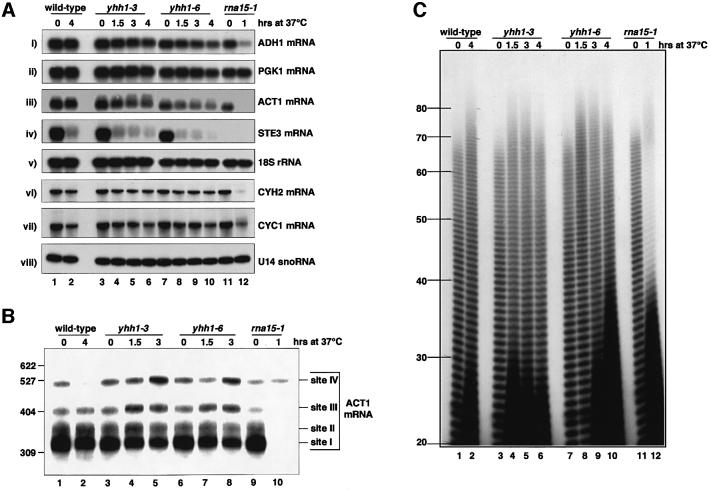
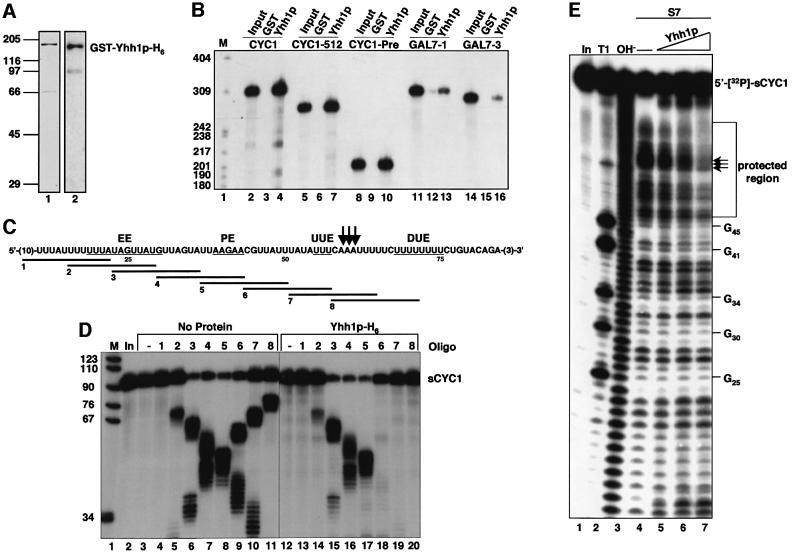
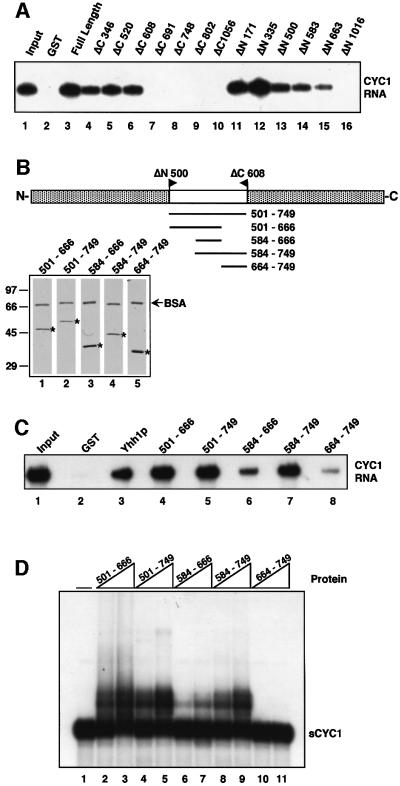
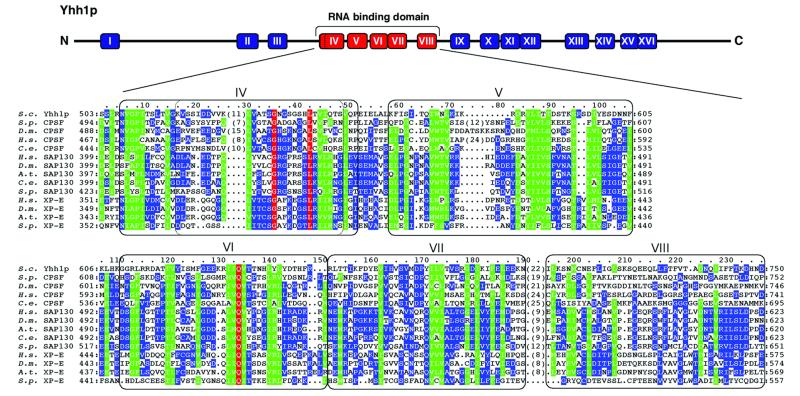
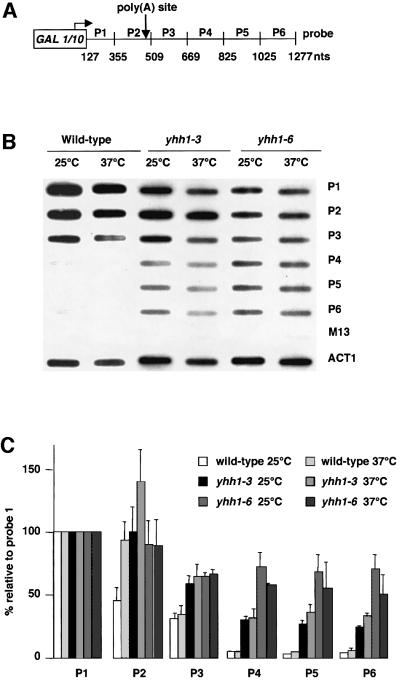
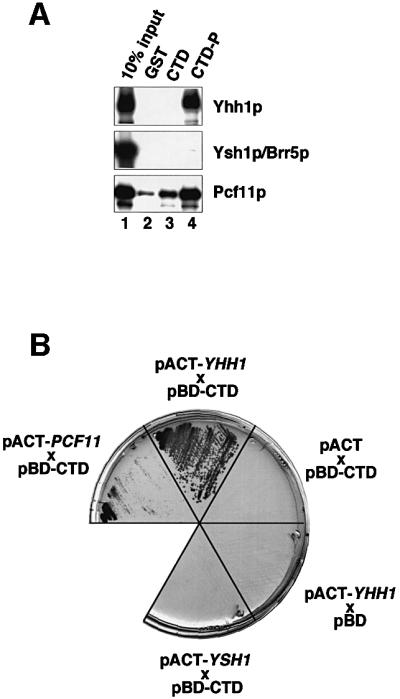
RNA polymerase II (pol II) transcription termination requires co-transcriptional recognition of a functional polyadenylation signal, but the molecular mechanisms that transduce this signal to pol II remain unclear. We show that Yhh1p/Cft1p, the yeast homologue of the mammalian AAUAAA interacting protein CPSF 160, is an RNA-binding protein and provide evidence that it participates in poly(A) site recognition. Interestingly, RNA binding is mediated by a central domain composed of predicted beta-propeller-forming repeats, which occurs in proteins of diverse cellular functions. We also found that Yhh1p/Cft1p bound specifically to the phosphorylated C-terminal domain (CTD) of pol II in vitro and in a two-hybrid test in vivo. Furthermore, transcriptional run-on analysis demonstrated that yhh1 mutants were defective in transcription termination, suggesting that Yhh1p/Cft1p functions in the coupling of transcription and 3'-end formation. We propose that direct interactions of Yhh1p/Cft1p with both the RNA transcript and the CTD are required to communicate poly(A) site recognition to elongating pol II to initiate transcription termination.
Figures
Similar articles
-
Cleavage/polyadenylation factor IA associates with the carboxyl-terminal domain of RNA polymerase II in Saccharomyces cerevisiae.Proc Natl Acad Sci U S A. 2001 Jan 16;98(2):445-50. doi: 10.1073/pnas.98.2.445. Epub 2001 Jan 9. Proc Natl Acad Sci U S A. 2001. PMID: 11149954 Free PMC article.
-
RNA polymerase II mutations conferring defects in poly(A) site cleavage and termination in Saccharomyces cerevisiae.G3 (Bethesda). 2013 Feb;3(2):167-80. doi: 10.1534/g3.112.004531. Epub 2013 Feb 1. G3 (Bethesda). 2013. PMID: 23390594 Free PMC article.
-
The role of the yeast cleavage and polyadenylation factor subunit Ydh1p/Cft2p in pre-mRNA 3'-end formation.Nucleic Acids Res. 2003 Jul 15;31(14):3936-45. doi: 10.1093/nar/gkg478. Nucleic Acids Res. 2003. PMID: 12853609 Free PMC article.
-
A history of poly A sequences: from formation to factors to function.Prog Nucleic Acid Res Mol Biol. 2002;71:285-389. doi: 10.1016/s0079-6603(02)71046-5. Prog Nucleic Acid Res Mol Biol. 2002. PMID: 12102557 Review.
-
[C-terminal domain (CTD) of the subunit Rpb1 of nuclear RNA polymerase II and its role in the transcription cycle].Mol Biol (Mosk). 2007 May-Jun;41(3):433-49. Mol Biol (Mosk). 2007. PMID: 17685222 Review. Russian.
Cited by
-
DNA damage induces targeted, genome-wide variation of poly(A) sites in budding yeast.Genome Res. 2013 Oct;23(10):1690-703. doi: 10.1101/gr.144964.112. Epub 2013 Jun 20. Genome Res. 2013. PMID: 23788651 Free PMC article.
-
Independent functions of yeast Pcf11p in pre-mRNA 3' end processing and in transcription termination.EMBO J. 2003 May 1;22(9):2167-77. doi: 10.1093/emboj/cdg200. EMBO J. 2003. PMID: 12727883 Free PMC article.
-
Unique features of plant cleavage and polyadenylation specificity factor revealed by proteomic studies.Plant Physiol. 2009 Nov;151(3):1546-56. doi: 10.1104/pp.109.142729. Epub 2009 Sep 11. Plant Physiol. 2009. PMID: 19748916 Free PMC article.
-
Cordycepin interferes with 3' end formation in yeast independently of its potential to terminate RNA chain elongation.RNA. 2009 May;15(5):837-49. doi: 10.1261/rna.1458909. Epub 2009 Mar 26. RNA. 2009. PMID: 19324962 Free PMC article.
-
Structural biology of poly(A) site definition.Wiley Interdiscip Rev RNA. 2011 Sep-Oct;2(5):732-47. doi: 10.1002/wrna.88. Epub 2011 Apr 27. Wiley Interdiscip Rev RNA. 2011. PMID: 21823232 Free PMC article. Review.
References
-
- Aranda A. and Proudfoot,N. (2001) Transcriptional termination factors for RNA polymerase II in yeast. Mol. Cell., 7, 1003–1011. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
