

Mechanisms and rates of bacterial colonization of sinking aggregates
- PMID: 12147501
- PMCID: PMC124032
- DOI: 10.1128/AEM.68.8.3996-4006.2002
Mechanisms and rates of bacterial colonization of sinking aggregates
Abstract
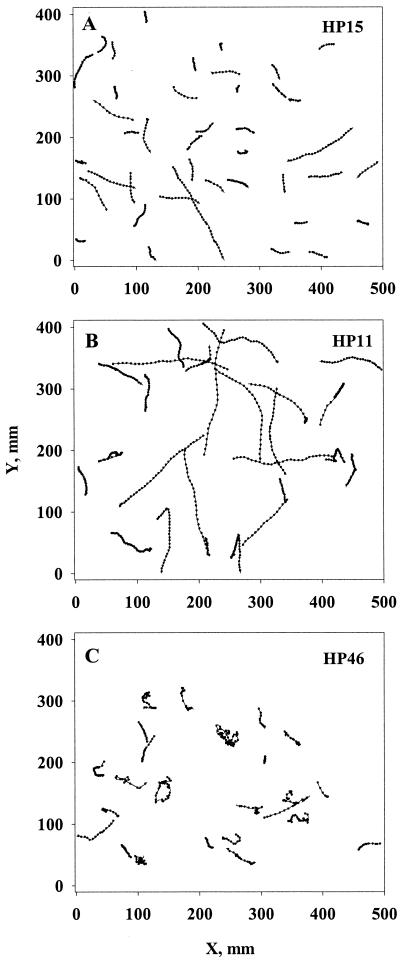
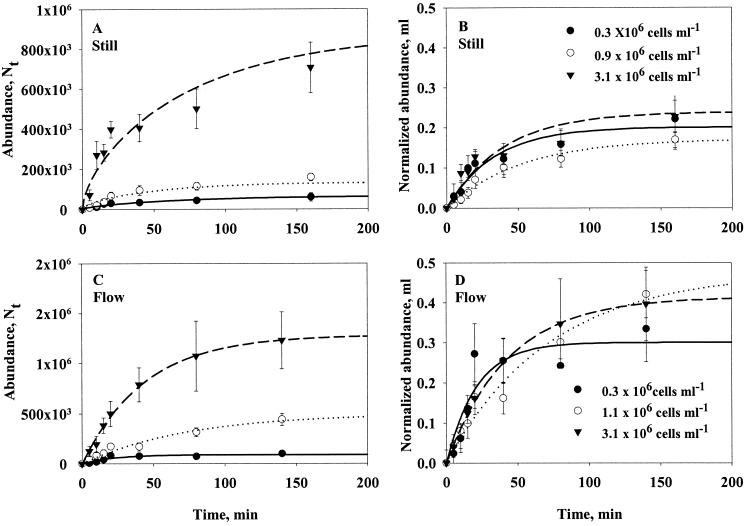
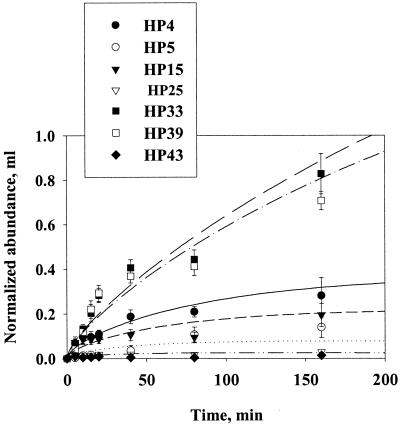
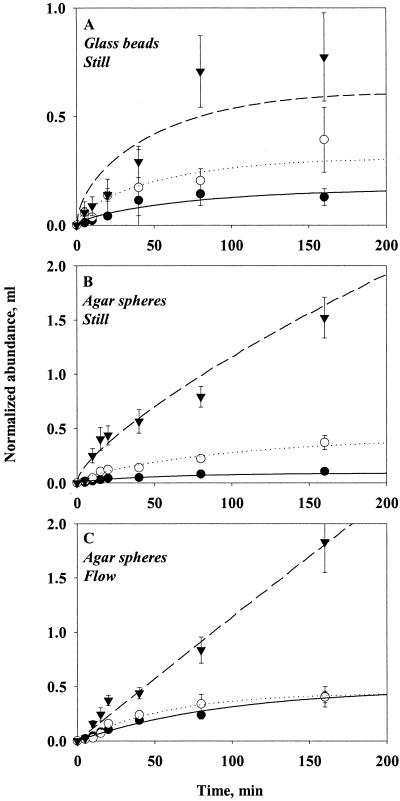
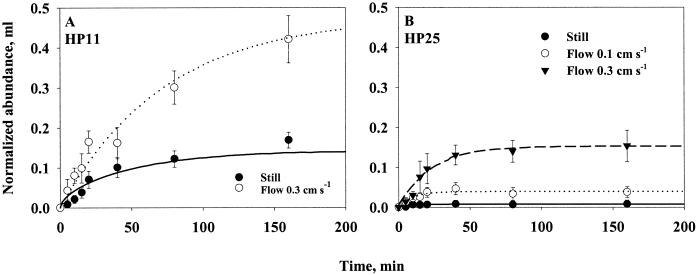
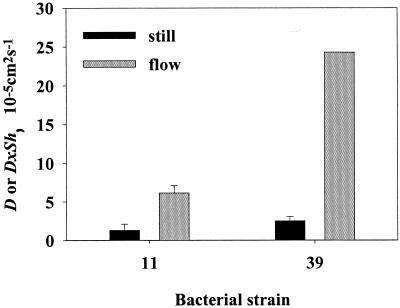
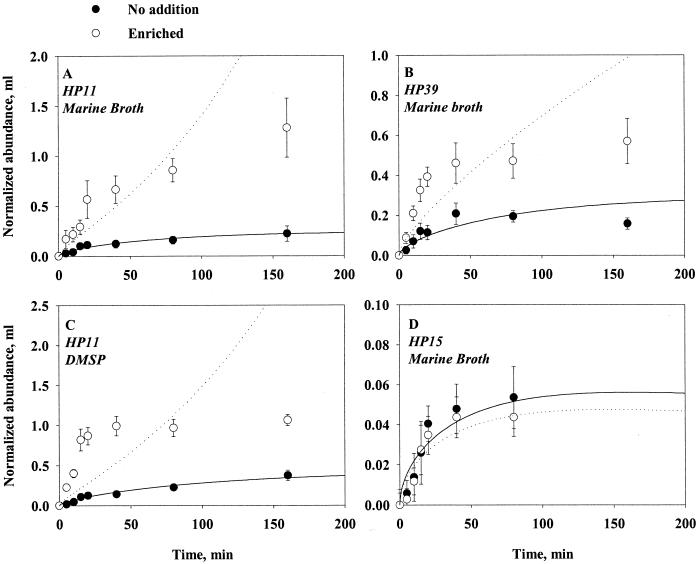
Quantifying the rate at which bacteria colonize aggregates is a key to understanding microbial turnover of aggregates. We used encounter models based on random walk and advection-diffusion considerations to predict colonization rates from the bacteria's motility patterns (swimming speed, tumbling frequency, and turn angles) and the hydrodynamic environment (stationary versus sinking aggregates). We then experimentally tested the models with 10 strains of bacteria isolated from marine particles: two strains were nonmotile; the rest were swimming at 20 to 60 microm s(-1) with different tumbling frequency (0 to 2 s(-1)). The rates at which these bacteria colonized artificial aggregates (stationary and sinking) largely agreed with model predictions. We report several findings. (i) Motile bacteria rapidly colonize aggregates, whereas nonmotile bacteria do not. (ii) Flow enhances colonization rates. (iii) Tumbling strains colonize aggregates enriched with organic substrates faster than unenriched aggregates, while a nontumbling strain did not. (iv) Once on the aggregates, the bacteria may detach and typical residence time is about 3 h. Thus, there is a rapid exchange between attached and free bacteria. (v) With the motility patterns observed, freely swimming bacteria will encounter an aggregate in <1 day at typical upper-ocean aggregate concentrations. This is faster than even starving bacteria burn up their reserves, and bacteria may therefore rely solely on aggregates for food. (vi) The net result of colonization and detachment leads to a predicted equilibrium abundance of attached bacteria as a function of aggregate size, which is markedly different from field observations. This discrepancy suggests that inter- and intraspecific interactions among bacteria and between bacteria and their predators may be more important than colonization in governing the population dynamics of bacteria on natural aggregates.
Figures




References
-
- Alldredge, A. L., and Y. Cohen. 1987. Can microscale chemical patches persist in the sea? Microelectrode study of marine snow and fecal pellets. Science 235:689-691. - PubMed
-
- Alldredge, A. L., J. J. Cole, and D. A. Caron. 1986. Production of heterotrophic bacteria inhabiting macroscopic aggregates (marine snow) from surface waters. Limnol. Oceanogr. 31:68-78.
-
- Alldredge, A. L., and C. Gotschalk. 1988. In situ settling behavior of marine snow. Limnol. Oceanogr. 33:339-351.
-
- Alldredge, A. L., and C. Gotschalk. 1990. The relative contribution of marine snow of different origin to biological processes in coastal waters. Cont. Shelf Res. 10:41-58.
-
- Alldredge, A. L., and M. Silver. 1988. Characteristics, dynamics, and significance of marine snow. Prog. Oceanogr. 20:41-58.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
