

Clinical resistance to the kinase inhibitor STI-571 in chronic myeloid leukemia by mutation of Tyr-253 in the Abl kinase domain P-loop
- PMID: 12149456
- PMCID: PMC125018
- DOI: 10.1073/pnas.162140299
Clinical resistance to the kinase inhibitor STI-571 in chronic myeloid leukemia by mutation of Tyr-253 in the Abl kinase domain P-loop
Abstract
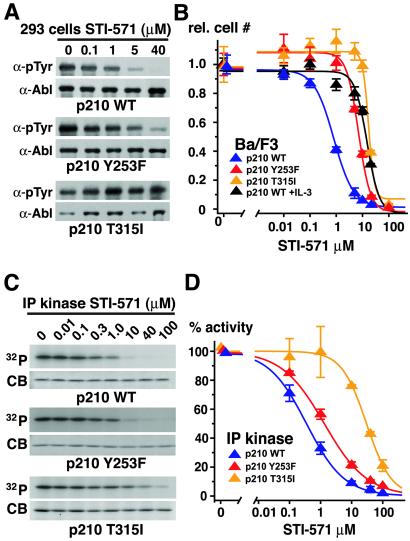
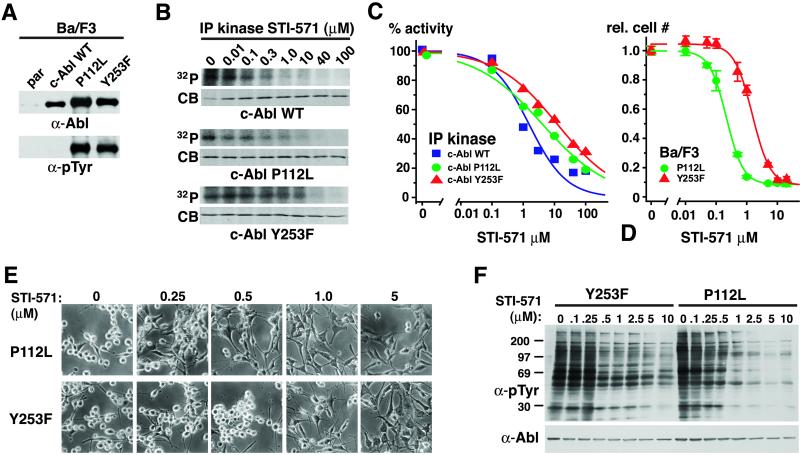
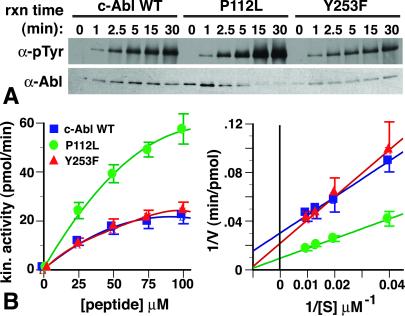
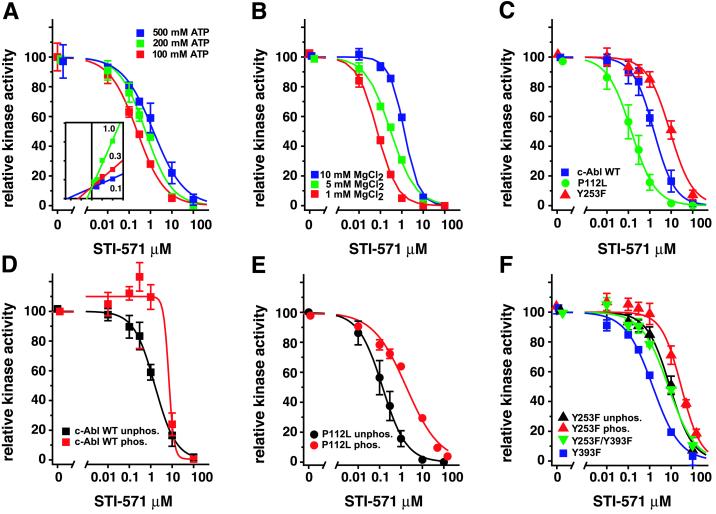
The Abl tyrosine kinase inhibitor STI-571 is effective therapy for stable phase chronic myeloid leukemia (CML) patients, but the majority of CML blast-crisis patients that respond to STI-571 relapse because of reactivation of Bcr-Abl signaling. Mutations of Thr-315 in the Abl kinase domain to Ile (T315I) were previously described in STI-571-resistant patients and likely cause resistance from steric interference with drug binding. Here we identify mutations of Tyr-253 in the nucleotide-binding (P) loop of the Abl kinase domain to Phe or His in patients with advanced CML and acquired STI-571 resistance. Bcr-Abl Y253F demonstrated intermediate resistance to STI-571 in vitro and in vivo when compared with Bcr-Abl T315I. The response of Abl proteins to STI-571 was influenced by the regulatory state of the kinase and by tyrosine phosphorylation. The sensitivity of purified c-Abl to STI-571 was increased by a dysregulating mutation (P112L) in the Src homology 3 domain of Abl but decreased by phosphorylation at the regulatory Tyr-393. In contrast, the Y253F mutation dysregulated c-Abl and conferred intrinsic but not absolute resistance to STI-571 that was independent of Tyr-393 phosphorylation. The Abl P-loop is a second target for mutations that confer resistance to STI-571 in advanced CML, and the Y253F mutation may impair the induced-fit interaction of STI-571 with the Abl catalytic domain rather than sterically blocking binding of the drug. Because clinical resistance induced by the Y253F mutation might be overcome by dose escalation of STI-571, molecular genotyping of STI-571-resistant patients may provide information useful for rational therapeutic management.
Figures
Similar articles
-
Molecular and chromosomal mechanisms of resistance to imatinib (STI571) therapy.Leukemia. 2002 Nov;16(11):2190-6. doi: 10.1038/sj.leu.2402741. Leukemia. 2002. PMID: 12399961 Clinical Trial.
-
BCR-ABL kinase domain mutations in tyrosine kinase inhibitors-naïve and -exposed Southeast Asian chronic myeloid leukemia patients.Exp Mol Pathol. 2012 Apr;92(2):259-65. doi: 10.1016/j.yexmp.2012.01.007. Epub 2012 Jan 31. Exp Mol Pathol. 2012. PMID: 22314255
-
Analysis of ABL kinase domain mutations conferring resistance to tyrosine kinase inhibitors in chronic myeloid leukemia cases from India.Clin Lab. 2011;57(7-8):619-23. Clin Lab. 2011. PMID: 21888027
-
Mechanisms and implications of imatinib resistance mutations in BCR-ABL.Curr Opin Hematol. 2004 Jan;11(1):35-43. doi: 10.1097/00062752-200401000-00006. Curr Opin Hematol. 2004. PMID: 14676625 Review.
-
Imatinib (STI571) resistance in chronic myelogenous leukemia: molecular basis of the underlying mechanisms and potential strategies for treatment.Mini Rev Med Chem. 2004 Mar;4(3):285-99. doi: 10.2174/1389557043487321. Mini Rev Med Chem. 2004. PMID: 15032675 Review.
Cited by
-
Phase 1 dose-finding study of rebastinib (DCC-2036) in patients with relapsed chronic myeloid leukemia and acute myeloid leukemia.Haematologica. 2017 Mar;102(3):519-528. doi: 10.3324/haematol.2016.152710. Epub 2016 Dec 7. Haematologica. 2017. PMID: 27927766 Free PMC article. Clinical Trial.
-
Blockade of Y177 and Nuclear Translocation of Bcr-Abl Inhibits Proliferation and Promotes Apoptosis in Chronic Myeloid Leukemia Cells.Int J Mol Sci. 2017 Mar 2;18(3):537. doi: 10.3390/ijms18030537. Int J Mol Sci. 2017. PMID: 28257089 Free PMC article.
-
Selective targeting of the inactive state of hematopoietic cell kinase (Hck) with a stable curcumin derivative.J Biol Chem. 2021 Jan-Jun;296:100449. doi: 10.1016/j.jbc.2021.100449. Epub 2021 Feb 20. J Biol Chem. 2021. PMID: 33617879 Free PMC article.
-
Controlling subcellular localization to alter function: Sending oncogenic Bcr-Abl to the nucleus causes apoptosis.J Control Release. 2009 Dec 16;140(3):245-9. doi: 10.1016/j.jconrel.2009.06.026. Epub 2009 Jul 1. J Control Release. 2009. PMID: 19576252 Free PMC article.
-
Mechanisms of drug resistance in kinases.Expert Opin Investig Drugs. 2011 Feb;20(2):153-208. doi: 10.1517/13543784.2011.546344. Expert Opin Investig Drugs. 2011. PMID: 21235428 Free PMC article. Review.
References
-
- Buchdunger E., Zimmermann, J., Mett, H., Meyer, T., Müller, M., Druker, B. J. & Lydon, N. B. (1996) Cancer Res. 56, 100-104. - PubMed
-
- Druker B. J., Tamura, S., Buchdunger, E., Ohno, S., Segal, G. M., Fanning, S., Zimmermann, J. & Lydon, N. B. (1996) Nat. Med. 2, 561-566. - PubMed
-
- Druker B. J., Talpaz, M., Resta, D. J., Peng, B., Buchdunger, E., Ford, J. M., Lydon, N. B., Kantarjian, H., Capdeville, R., Ohno-Jones, S. & Sawyers, C. L. (2001) N. Engl. J. Med. 344, 1031-1037. - PubMed
-
- Druker B. J., Sawyers, C. L., Kantarjian, H., Resta, D. J., Reese, S. F., Ford, J. M., Capdeville, R. & Talpaz, M. (2001) N. Engl. J. Med. 344, 1038-1042. - PubMed
-
- Gambacorti-Passerini C., Barni, R., le Coutre, P., Zucchetti, M., Cabrita, G., Cleris, L., Rossi, F., Gianazza, E., Brueggen, J., Cozens, R., et al. (2000) J. Natl. Cancer Inst. 92, 1641-1650. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
