

Supply-demand balance and metabolic scaling
- PMID: 12149461
- PMCID: PMC124956
- DOI: 10.1073/pnas.162216899
Supply-demand balance and metabolic scaling
Abstract
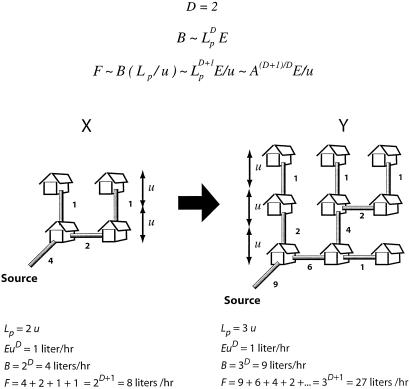
It is widely accepted that metabolic rates scale across species approximately as the 3/4 power of mass in most if not all groups of organisms. Metabolic demand per unit mass thus decreases as body mass increases. Metabolic rates reflect both the ability of the organism's transport system to deliver metabolites to the tissues and the rate at which the tissues use them. We show that the ubiquitous 3/4 power law for interspecific metabolic scaling arises from simple, general geometric properties of transportation networks constrained to function in biological organisms. The 3/4 exponent and other observed scaling relationships follow when mass-specific metabolic demands match the changing delivery capacities of the network at different body sizes. Deviation from the 3/4 exponent suggests either inefficiency or compensating physiological mechanisms. Our conclusions are based on general arguments incorporating the minimum of biological detail and should therefore apply to the widest range of organisms.
Figures
Comment in
-
Comment on "Revising the distributive networks models of West, Brown and Enquist (1997) and Banavar, Maritan and Rinaldo (1999): Metabolic inequity of living tissues provides clues for the observed allometric scaling rules" by Makarieva, Gorshkov and Li.J Theor Biol. 2006 Apr 7;239(3):391-3. doi: 10.1016/j.jtbi.2005.08.023. Epub 2005 Sep 28. J Theor Biol. 2006. PMID: 16197962 No abstract available.
References
-
- McMahon T. A. & Bonner, J. T., (1983) On Size and Life (Freeman, New York).
-
- Bonner J. T., (1983) The Evolution of Complexity by Means of Natural Selection (Princeton Univ. Press, Princeton).
-
- Peters R. H., (1983) The Ecological Implications of Body Size (Cambridge Univ. Press, Cambridge, U.K.).
-
- Schmidt-Nielsen K., (1984) Scaling: Why Is Animal Size So Important? (Cambridge Univ. Press, Cambridge, U.K.).
-
- Calder W. A., (1984) Size, Function, and Life History (Harvard Univ. Press, Cambridge, MA).
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
