

Closure of the Venus flytrap module of mGlu8 receptor and the activation process: Insights from mutations converting antagonists into agonists
- PMID: 12151600
- PMCID: PMC123216
- DOI: 10.1073/pnas.162138699
Closure of the Venus flytrap module of mGlu8 receptor and the activation process: Insights from mutations converting antagonists into agonists
Abstract
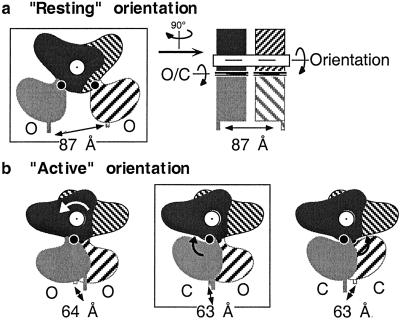
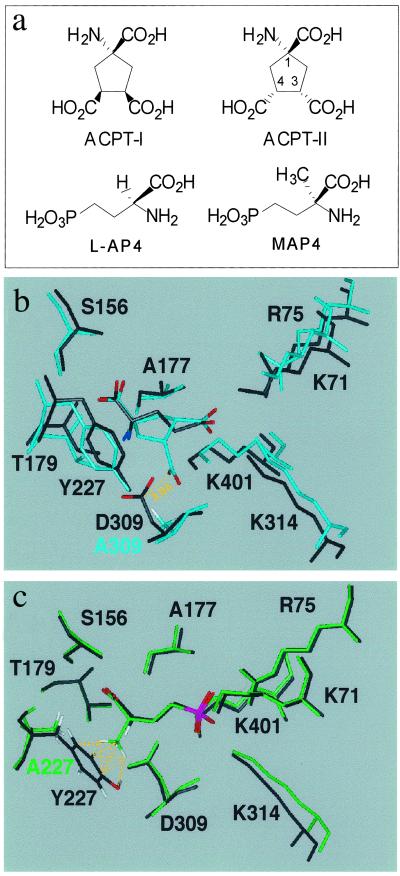
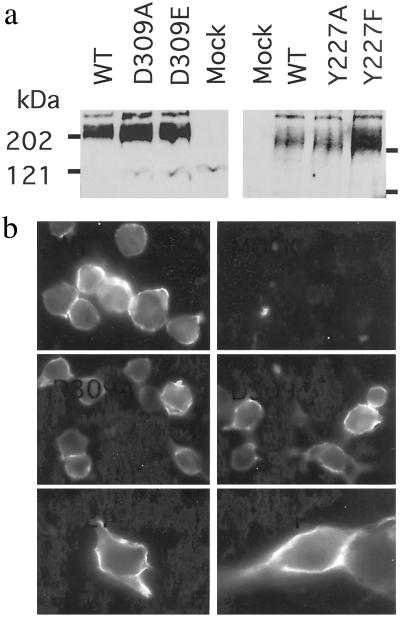
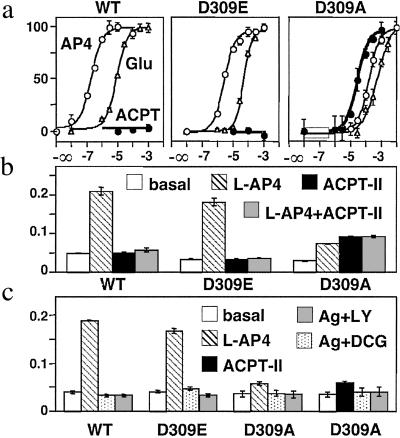
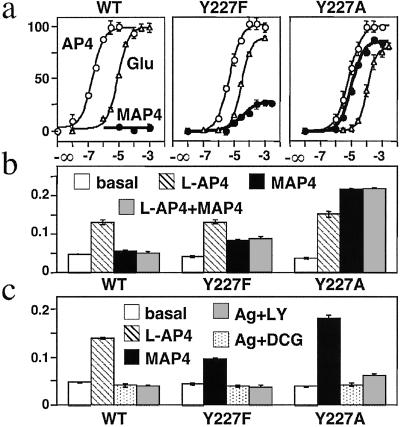
Ca2+, pheromones, sweet taste compounds, and the main neurotransmitters glutamate and gamma-aminobutyric acid activate G protein-coupled receptors (GPCRs) that constitute the GPCR family 3. These receptors are dimers, and each subunit has a large extracellular domain called a Venus flytrap module (VFTM), where agonists bind. This module is connected to a heptahelical domain that activates G proteins. Recently, the structure of the dimer of mGlu1 VFTMs revealed two important conformational changes resulting from glutamate binding. First, agonists can stabilize a closed state of at least one VFTM in the dimer. Second, the relative orientation of the two VFTMs in the dimer is different in the presence of glutamate, such that their C-terminal ends (which are connected to the G protein-activating heptahelical domain) become closer by more than 20 A. This latter change in orientation has been proposed to play a key role in receptor activation. To elucidate the respective role of VFTM closure and the change in orientation of the VFTMs in family 3 GPCR activation, we analyzed the mechanism of action of the mGlu8 receptor antagonists ACPT-II and MAP4. Molecular modeling studies suggest that these two compounds prevent the closure of the mGlu8 VFTM because of ionic and steric hindrance, respectively. We show here that the replacement of the residues responsible for these hindrances (Asp-309 and Tyr-227, respectively) by Ala allows ACPT-II or MAP4 to fully activate the receptors. These data are consistent with the requirement of the VFTM closure for family 3 GPCR activation.
Figures
References
-
- De Blasi A., Conn, P. J., Pin, J.-P. & Nicoletti, F. (2001) Trends Pharmacol. Sci. 22, 114-120. - PubMed
-
- Takahashi K., Tsuchida, K., Tanabe, Y., Masu, M. & Nakanishi, S. (1993) J. Biol. Chem. 268, 19341-19345. - PubMed
-
- O'Hara P. J., Sheppard, P. O., Thøgersen, H., Venezia, D., Haldeman, B. A., McGrane, V., Houamed, K. M., Thomsen, C., Gilbert, T. L. & Mulvihill, E. R. (1993) Neuron 11, 41-52. - PubMed
-
- Costantino G. & Pellicciari, R. (1996) J. Med. Chem. 39, 3998-4006. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
