

A new Groucho TLE4 protein may regulate the repressive activity of Pax5 in human B lymphocytes
- PMID: 12153506
- PMCID: PMC1782747
- DOI: 10.1046/j.1365-2567.2002.01456.x
A new Groucho TLE4 protein may regulate the repressive activity of Pax5 in human B lymphocytes
Abstract
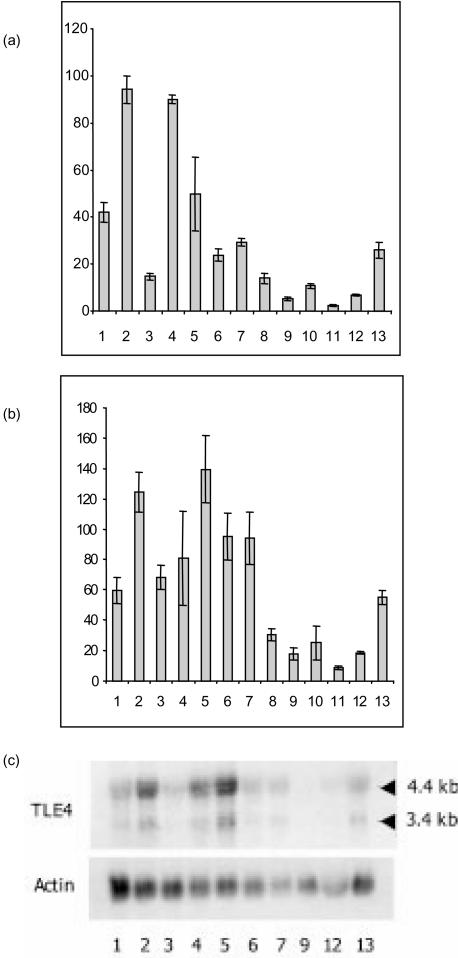
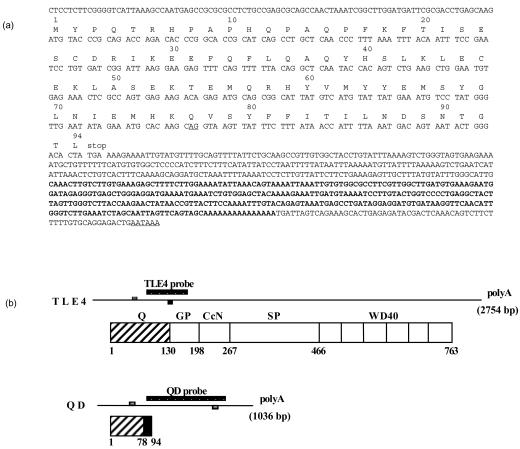
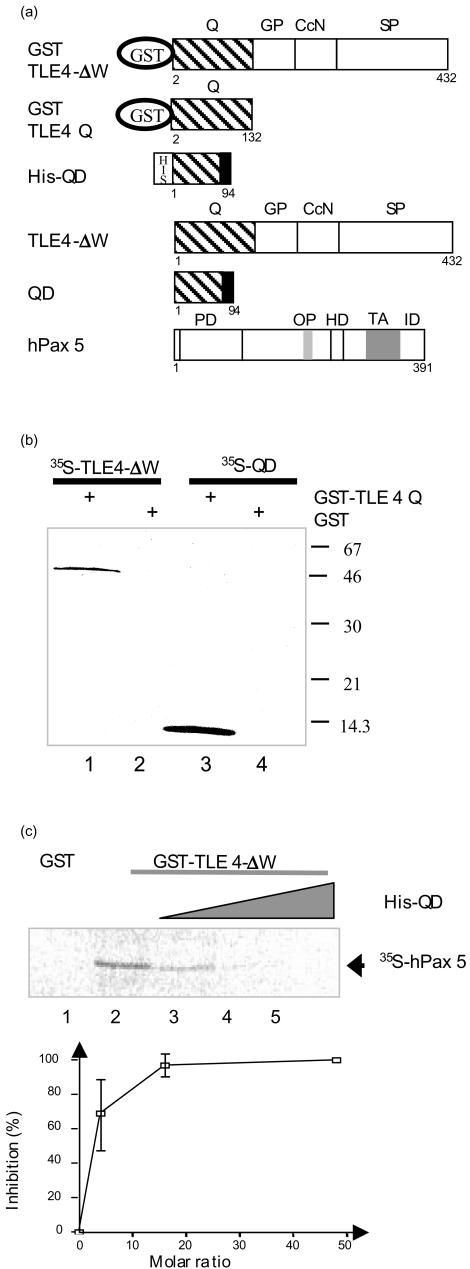
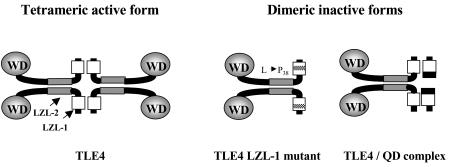
During mouse B-cell development, Pax5 is an essential transcription factor that acts as an activator of B-cell-specific genes and as a repressor of alternative lineage fates. The repressive function is mediated by the recruitment of members of the Groucho co-repressor family. Using an RNA display approach, we have isolated a transcript, called QD, specifically expressed in human pro-B and pre-B cells, which is derived from the human Groucho TLE4 gene. The QD transcript contains the first four TLE4 exons and an intronic sequence 3' of exon 4, demonstrating that QD is a splice variant of TLE4. The putative resulting protein of 94 amino acids corresponds to approximately half of an N-terminal tetramerization domain. We also show specific expression of TLE4 transcripts in human B cells and of TLE4 proteins in B-cell nuclei. Moreover, we demonstrate that recombinant QD protein binds to the TLE4 Q domain and is able to abolish the TLE4/Pax5 interaction. Thus, QD could act as a negative regulator of TLE4 function, in early B-cell differentiation.
Figures


Similar articles
-
The germ cell nuclear proteins hnRNP G-T and RBMY activate a testis-specific exon.PLoS Genet. 2009 Nov;5(11):e1000707. doi: 10.1371/journal.pgen.1000707. Epub 2009 Nov 6. PLoS Genet. 2009. PMID: 19893608 Free PMC article.
-
Commitment to the B-lymphoid lineage depends on the transcription factor Pax5.Nature. 1999 Oct 7;401(6753):556-62. doi: 10.1038/44076. Nature. 1999. PMID: 10524622
-
Transcriptional repression by Pax5 (BSAP) through interaction with corepressors of the Groucho family.EMBO J. 2000 May 15;19(10):2292-303. doi: 10.1093/emboj/19.10.2292. EMBO J. 2000. PMID: 10811620 Free PMC article.
-
BSAP: a key regulator of B-cell development and differentiation.Immunol Today. 1995 Dec;16(12):564-9. doi: 10.1016/0167-5699(95)80078-6. Immunol Today. 1995. PMID: 8579748 Review.
-
Pax5 determines the identity of B cells from the beginning to the end of B-lymphopoiesis.Int Rev Immunol. 2001 Feb;20(1):65-82. doi: 10.3109/08830180109056723. Int Rev Immunol. 2001. PMID: 11342298 Review.
Cited by
-
Identification of novel variants in colorectal cancer families by high-throughput exome sequencing.Cancer Epidemiol Biomarkers Prev. 2013 Jul;22(7):1239-51. doi: 10.1158/1055-9965.EPI-12-1226. Epub 2013 May 1. Cancer Epidemiol Biomarkers Prev. 2013. PMID: 23637064 Free PMC article.
-
Hacker within! Ehrlichia chaffeensis Effector Driven Phagocyte Reprogramming Strategy.Front Cell Infect Microbiol. 2016 May 31;6:58. doi: 10.3389/fcimb.2016.00058. eCollection 2016. Front Cell Infect Microbiol. 2016. PMID: 27303657 Free PMC article. Review.
-
Genome-wide association study to identify novel loci associated with therapy-related myeloid leukemia susceptibility.Blood. 2009 May 28;113(22):5575-82. doi: 10.1182/blood-2008-10-183244. Epub 2009 Mar 18. Blood. 2009. PMID: 19299336 Free PMC article.
-
Downregulation of RUNX1 by RUNX3 requires the RUNX3 VWRPY sequence and is essential for Epstein-Barr virus-driven B-cell proliferation.J Virol. 2009 Jul;83(13):6909-16. doi: 10.1128/JVI.00216-09. Epub 2009 Apr 29. J Virol. 2009. PMID: 19403666 Free PMC article.
-
The Groucho/Transducin-like enhancer of split protein family in animal development.IUBMB Life. 2015 Jul;67(7):472-81. doi: 10.1002/iub.1395. Epub 2015 Jul 14. IUBMB Life. 2015. PMID: 26172616 Free PMC article. Review.
References
-
- Nutt SL, Urbanek P, Rolink A, Busslinger M. Essential functions of Pax5 (BSAP) in pro-B cell development: difference between fetal and adult B lymphopoiesis and reduced V-to-DJ recombination at the IgH locus. Genes Dev. 1997;11:476–91. - PubMed
-
- O'Riordan M, Grosschedl R. Coordinate regulation of B cell differentiation by the transcription factors EBF and E2A. Immunity. 1999;11:21–31. - PubMed
-
- Bain G, Robanus Maandag EC, te Riele HP, et al. Both E12 and E47 allow commitment to the B cell lineage. Immunity. 1997;6:145–54. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
