

Molecular mechanisms governing Pcdh-gamma gene expression: evidence for a multiple promoter and cis-alternative splicing model
- PMID: 12154121
- PMCID: PMC186422
- DOI: 10.1101/gad.1004802
Molecular mechanisms governing Pcdh-gamma gene expression: evidence for a multiple promoter and cis-alternative splicing model
Abstract
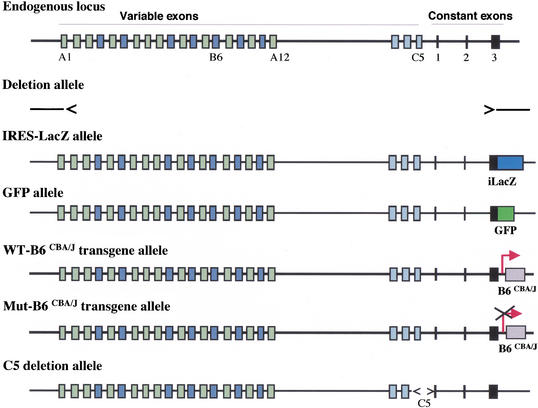
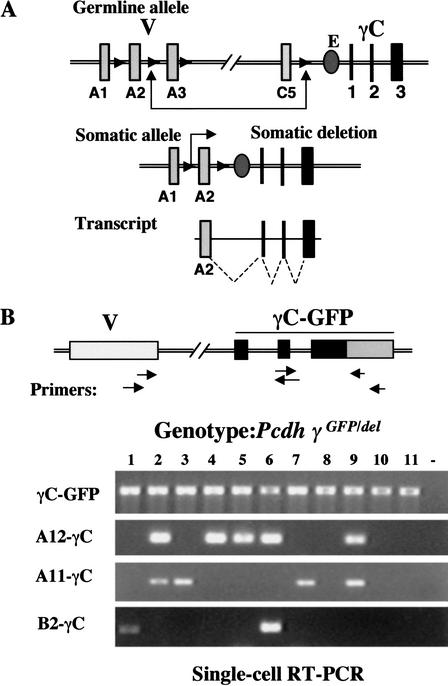
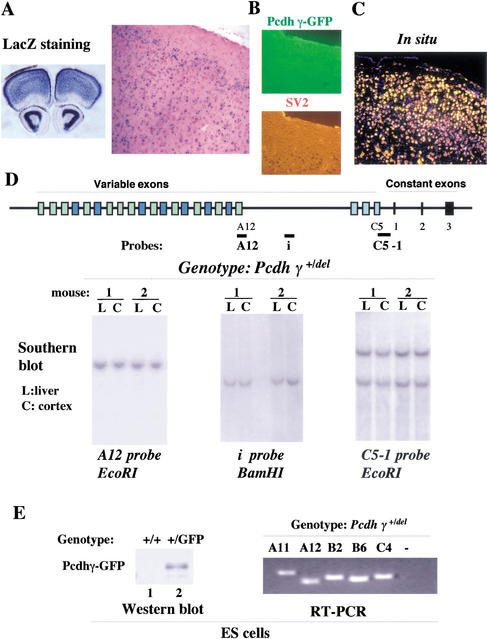
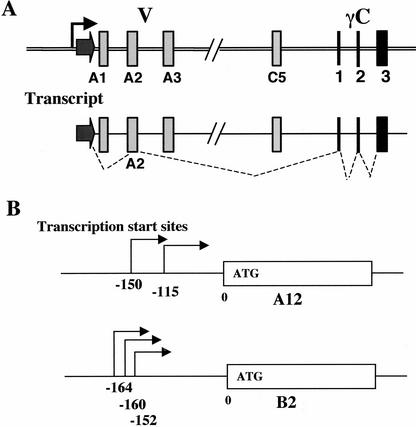
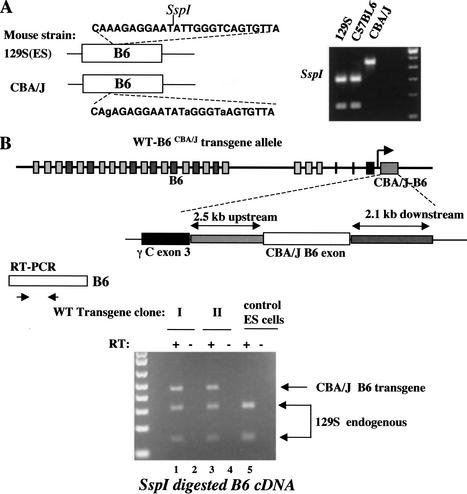
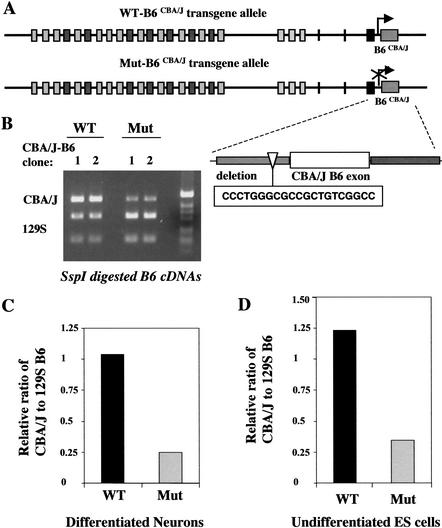
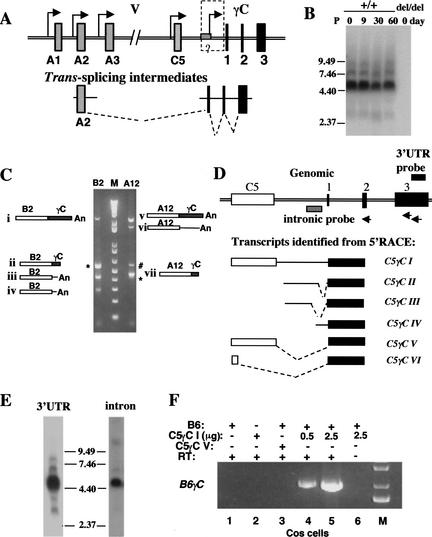
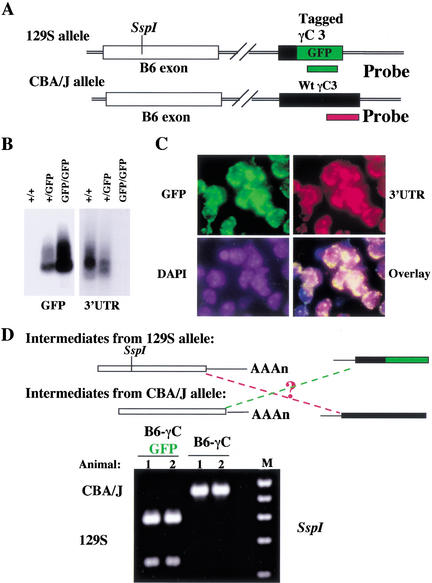
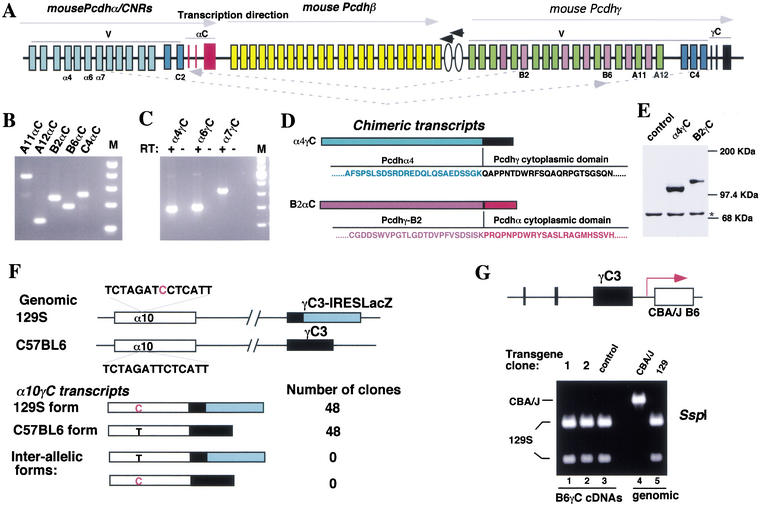
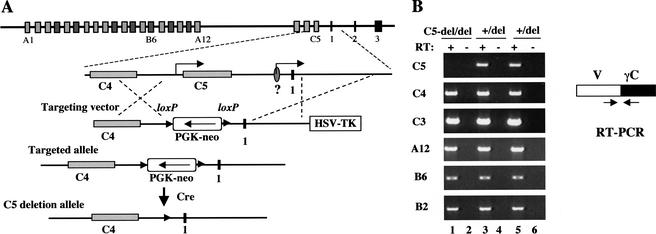
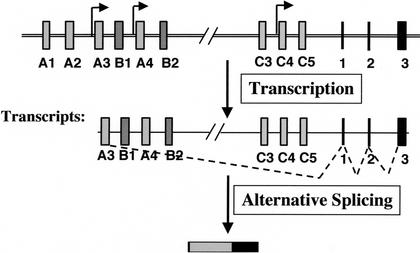
The genomic architecture of protocadherin (Pcdh) gene clusters is remarkably similar to that of the immunoglobulin and T cell receptor gene clusters, and can potentially provide significant molecular diversity. Pcdh genes are abundantly expressed in the central nervous system. These molecules are primary candidates for establishing specific neuronal connectivity. Despite the extensive analyses of the genomic structure of both human and mouse Pcdh gene clusters, the definitive molecular mechanisms that control Pcdh gene expression are still unknown. Four theories have been proposed, including (1) DNA recombination followed by cis-splicing, (2) single promoter and cis-alternative splicing, (3) multiple promoters and cis-alternative splicing, and (4) multiple promoters and trans-splicing. Using a combination of molecular and genetic analyses, we evaluated the four models at the Pcdh-gamma locus. Our analysis provides evidence that the transcription of individual Pcdh-gamma genes is under the control of a distinct but related promoter upstream of each Pcdh-gamma variable exon, and posttranscriptional processing of each Pcdh-gamma transcript is predominantly mediated through cis-alternative splicing.
Figures
References
-
- Agabian N. Trans splicing of nuclear pre-mRNAs. Cell. 1990;61:1157–1160. - PubMed
-
- Akopian AN, Okuse K, Souslova V, England S, Ogata N, Wood JN. Trans-splicing of a voltage-gated sodium channel is regulated by nerve growth factor. FEBS Lett. 1999;445:177–182. - PubMed
-
- Bain G, Kitchens D, Yao M, Huettner JE, Gottlieb DI. Embryonic stem cells express neuronal properties in vitro. Dev Biol. 1995;168:342–357. - PubMed
-
- Benson DL, Colman DR, Huntley GW. Molecules, maps and synapse specificity. Nat Rev Neurosci. 2001;2:899–909. - PubMed
-
- Bonen L. Trans-splicing of pre-mRNA in plants, animals, and protists. FASEB J. 1993;7:40–46. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases