

Differential chromatin structure within a tandem array 100 kb upstream of the maize b1 locus is associated with paramutation
- PMID: 12154122
- PMCID: PMC186425
- DOI: 10.1101/gad.1006702
Differential chromatin structure within a tandem array 100 kb upstream of the maize b1 locus is associated with paramutation
Abstract
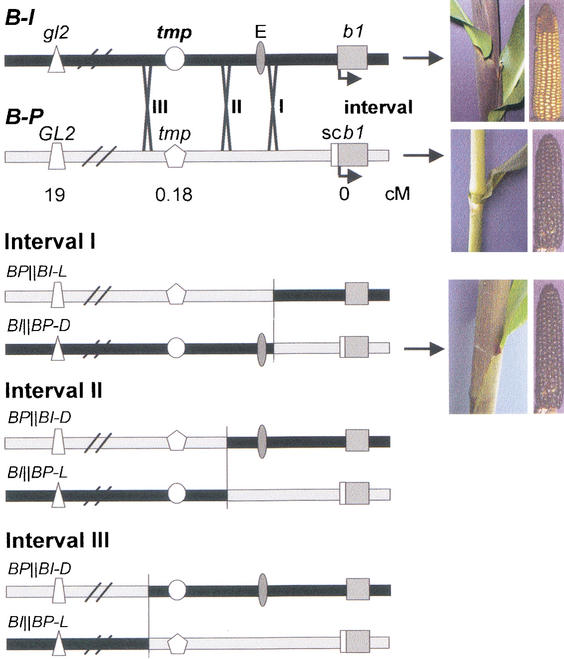
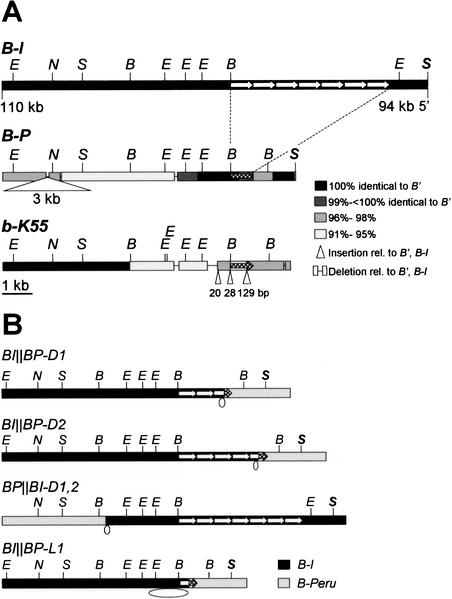
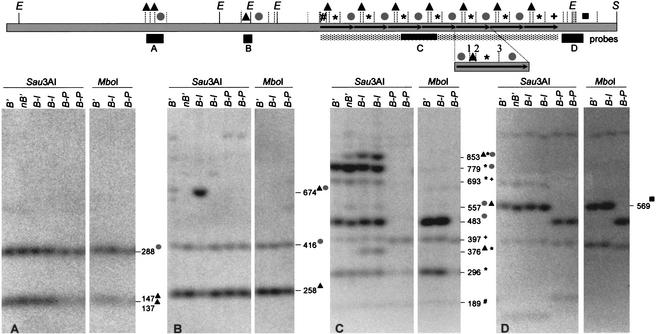
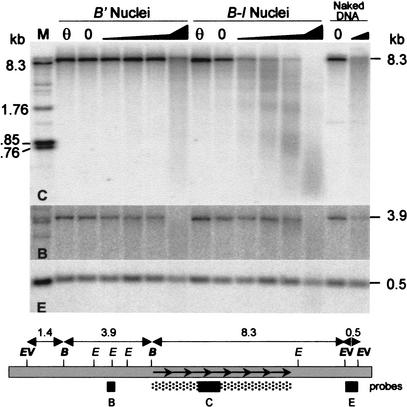
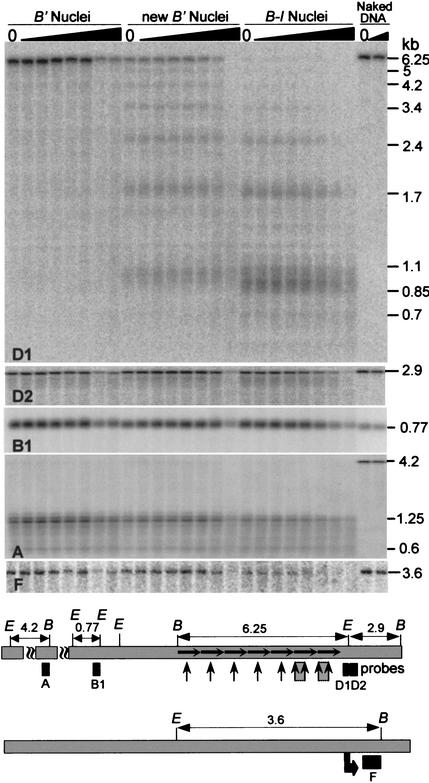
Recombination mapping defined a 6-kb region, 100 kb upstream of the transcription start site, that is required for B-I enhancer activity and paramutation-a stable, heritable change in transcription caused by allele interactions in maize (Zea mays). In this region, B-I and B' (the only b1 alleles that participate in paramutation) have seven tandem repeats of an 853-bp sequence otherwise unique in the genome; other alleles have one. Examination of recombinant alleles with different numbers of tandem repeats indicates that the repeats are required for both paramutation and enhancer function. The 6-kb region is identical in B-I and B', showing that epigenetic mechanisms mediate the stable silencing associated with paramutation. This is the first endogenous gene for which sequences required for paramutation have been defined and examined for methylation and chromatin structure. The tandem repeat sequences are more methylated in B-I (high expressing) relative to B' (low expressing), opposite of the typical correlation. Furthermore, the change in repeat methylation follows establishment of the B' epigenetic state. B-I has a more open chromatin structure in the repeats relative to B'. The nuclease hypersensitivity differences developmentally precede transcription, suggesting that the repeat chromatin structure could be the heritable imprint distinguishing the two transcription states.
Figures
References
-
- Barlow DP. Gametic imprinting in mammals. Science. 1995;270:1610–1613. - PubMed
-
- Bell AC, Felsenfeld G. Methylation of a CTCF-dependent boundary controls imprinted expression of the Igf2 gene. Nature. 2000;405:482–485. - PubMed
-
- Bell AC, West AG, Felsenfeld G. Insulators and boundaries: Versatile regulatory elements in the eukaryotic genome. Science. 2001;291:447–449. - PubMed
-
- Boumil RM, Lee JT. Forty years of decoding the silence in X-chromosome inactivation. Hum Mol Genet. 2001;10:2225–2232. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources