

Phytochrome mediates the external light signal to repress FT orthologs in photoperiodic flowering of rice
- PMID: 12154129
- PMCID: PMC186415
- DOI: 10.1101/gad.999202
Phytochrome mediates the external light signal to repress FT orthologs in photoperiodic flowering of rice
Abstract
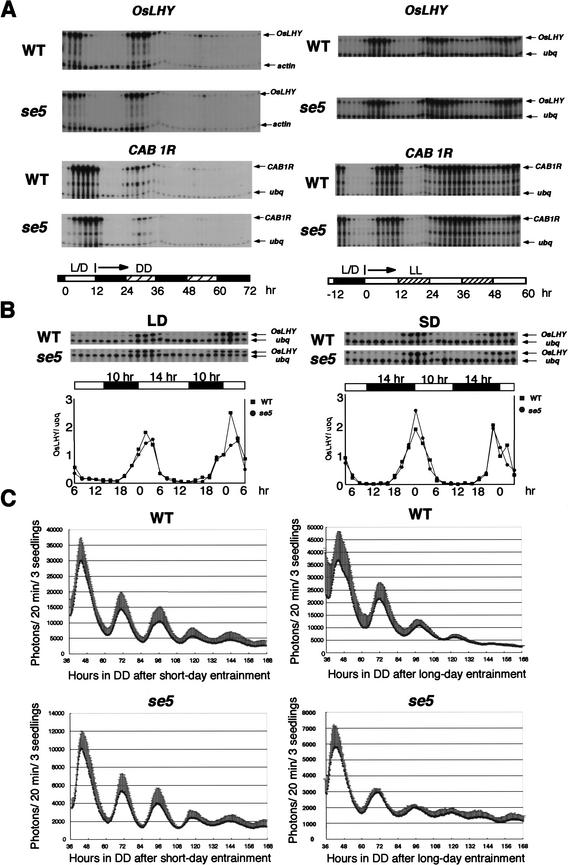
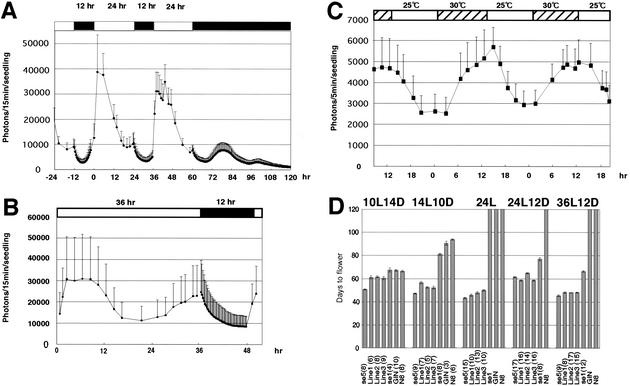
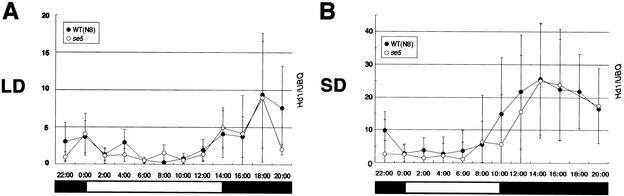
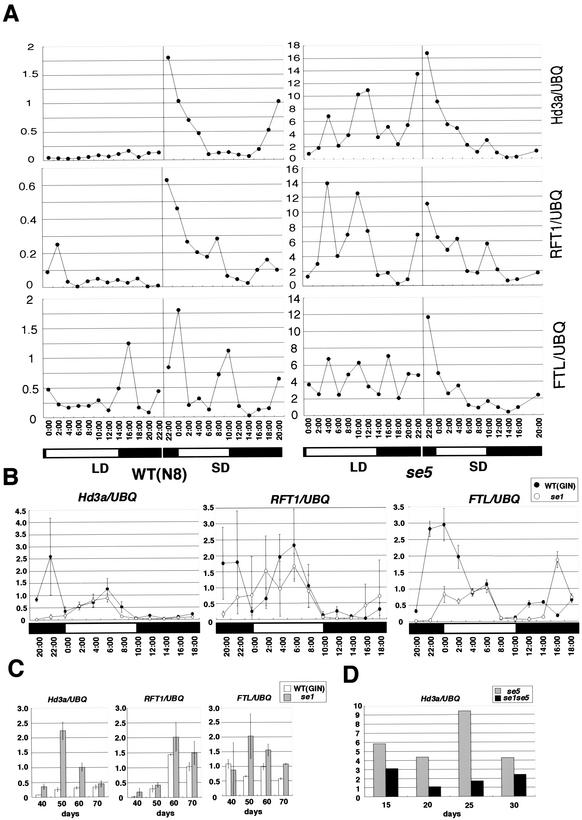
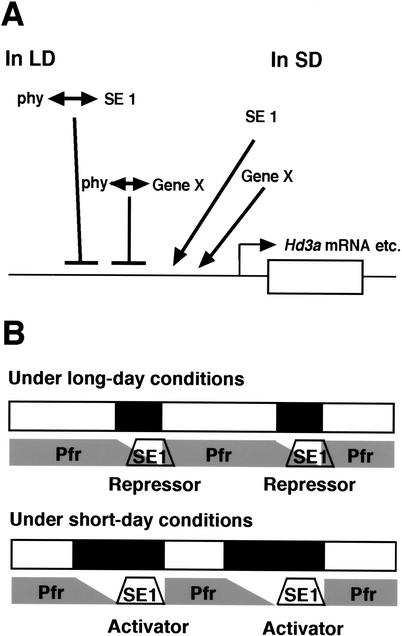
Phytochromes confer the photoperiodic control of flowering in rice (Oryza sativa), a short-day plant. To better understand the molecular mechanisms of day-length recognition, we examined the interaction between phytochrome signals and circadian clocks in photoperiodic-flowering mutants of rice. Monitoring behaviors of circadian clocks revealed that phase setting of circadian clocks is not affected either under short-day (SD) or under long-day (LD) conditions in a phytochrome-deficient mutant that shows an early-flowering phenotype with no photoperiodic response. Non-24-hr-light/dark-cycle experiments revealed that a rice counterpart gene of Arabidopsis CONSTANS (CO), named PHOTOPERIOD SENSITIVITY 1 (Heading date 1) [SE1 (Hd1)], functions as an output of circadian clocks. In addition, the phytochrome deficiency does not affect the diurnal mRNA expression of SE1 upon floral transition. Downstream floral switch genes were further identified with rice orthologs of Arabidopsis FLOWERING LOCUS T (FT). Our RT-PCR data indicate that phytochrome signals repress mRNA expression of FT orthologs, whereas SE1 can function to promote and suppress mRNA expression of the FT orthologs under SD and LD, respectively. This SE1 transcriptional activity may be posttranscriptionally regulated and may depend on the coincidence with Pfr phytochromes. We propose a model to explain how a short-day plant recognizes the day length in photoperiodic flowering.
Figures

References
-
- Alabadi D, Oyama T, Yanovsky MJ, Harmon FG, Mas P, Kay SA. Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science. 2001;293:880–883. - PubMed
-
- Bradley D, Ratcliffe O, Vincent C, Carpenter R, Coen E. Inflorescence commitment and architecture in Arabidopsis. Science. 1997;275:80–83. - PubMed
-
- Bünning E. Circadian rhythms and the time measurement in photoperiodism. Cold Spring Harbor Symp Quant Biol. 1960;25:249–256. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials