

Molecular characterization of preintegration latency in human immunodeficiency virus type 1 infection
- PMID: 12163571
- PMCID: PMC136977
- DOI: 10.1128/jvi.76.17.8518-8513.2002
Molecular characterization of preintegration latency in human immunodeficiency virus type 1 infection
Abstract
Most current evidence suggests that the infection of resting CD4(+) T cells by human immunodeficiency virus type 1 (HIV-1) is not productive due to partial or complete blocks in the viral life cycle at steps prior to integration of the viral genome into the host cell chromosome. However, stimulation of an infected resting T cell by antigen, cytokines, or microenvironmental factors can overcome these blocks and allow for the production of progeny virions. In this study, we sought to understand the structure and fate of the virus in unstimulated resting CD4(+) T cells. Using a novel linker-mediated PCR assay designed to detect and characterize linear unintegrated forms of the HIV-1 genome, we demonstrate that reverse transcription can proceed to completion following the infection of resting T cells, generating the substrate for the retroviral integration reaction. However, reverse transcription in resting T cells is far slower than in activated T cells, requiring 2 to 3 days to complete. The delay in completing reverse transcription may make the viral DNA genome more susceptible to competing decay processes. To explore the relationship between the formation of the linear viral genome and the stability of the preintegration state, we employed a recombinant HIV-1 virus expressing the enhanced green fluorescent protein to measure the rate at which HIV-1 decays in the preintegration state. Our results demonstrate that the preintegration state is labile and decays rapidly (half-life = 1 day) following the entry of HIV-1 into a resting T cell, with significant decay occurring during the slow process of reverse transcription.
Figures



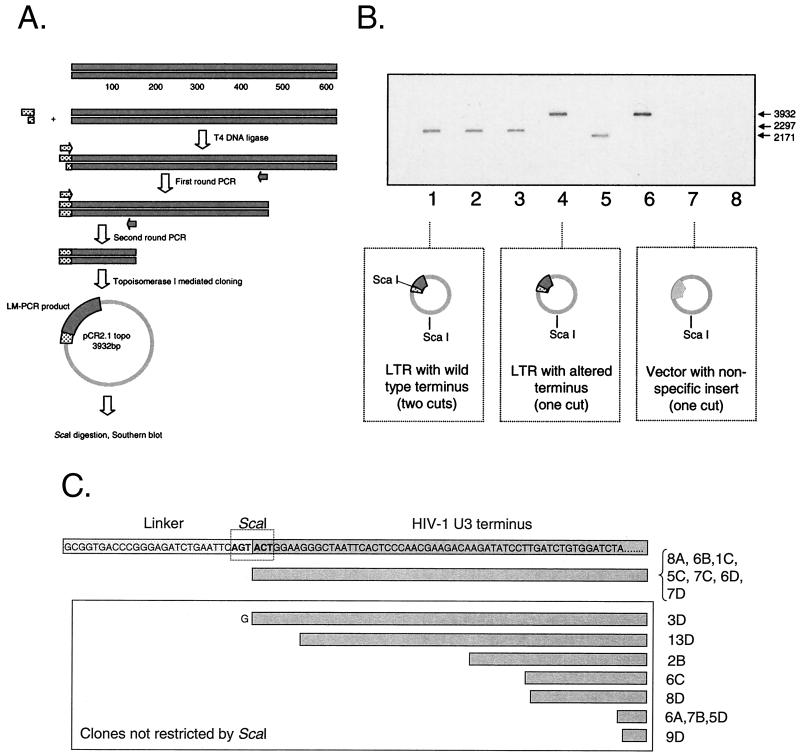
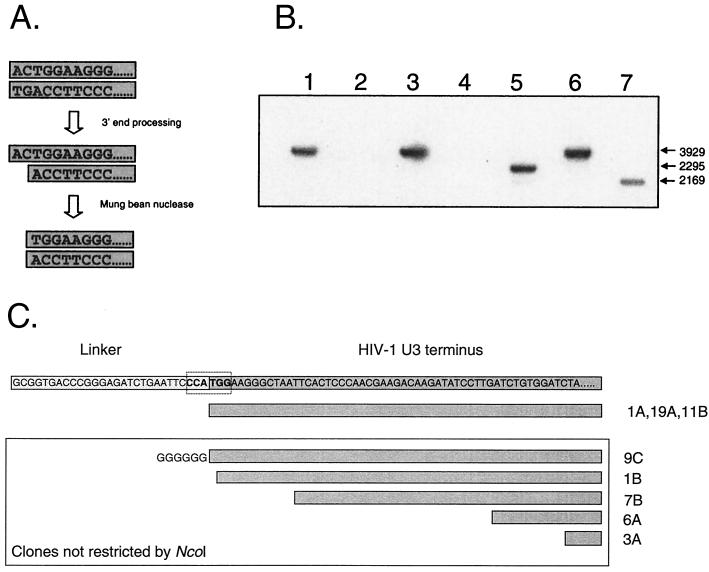
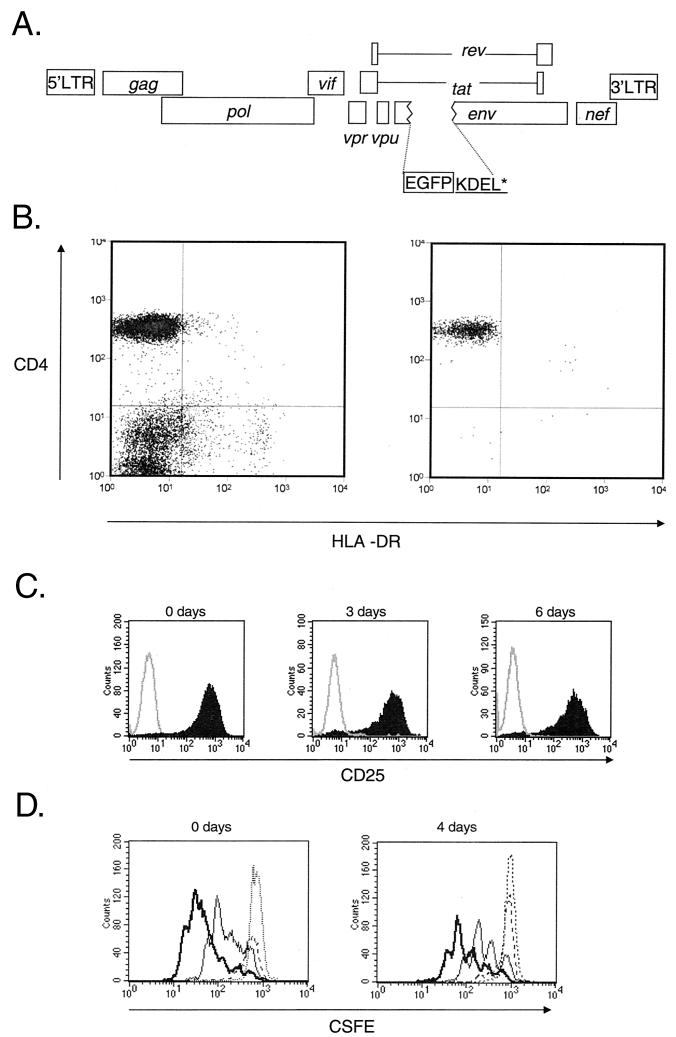
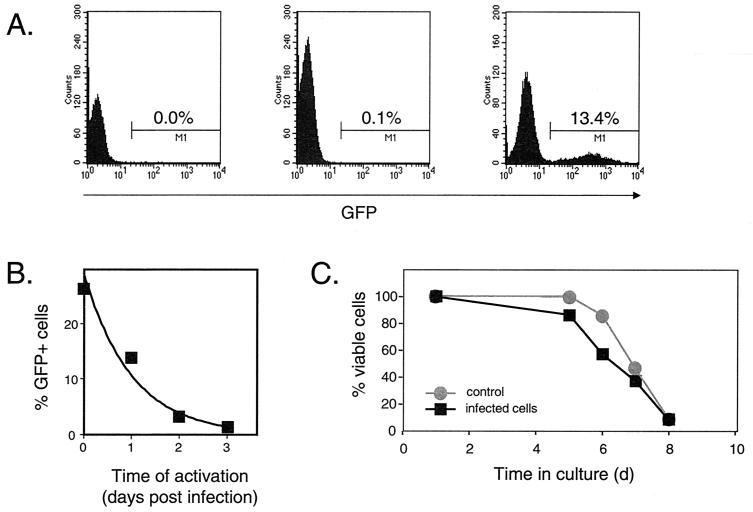

References
-
- Blankson, J. N., D. Finzi, T. C. Pierson, B. P. Sabundayo, K. Chadwick, J. B. Margolick, T. C. Quinn, and R. F. Siliciano. 2000. Biphasic decay of latently infected CD4+ T cells in acute human immunodeficiency virus type 1 infection. J. Infect. Dis. 182:1636-1642. - PubMed
-
- Chun, T. W., L. Carruth, D. Finzi, X. Shen, J. A. DiGiuseppe, H. Taylor, M. Hermankova, K. Chadwick, J. Margolick, T. C. Quinn, Y. H. Kuo, R. Brookmeyer, M. A. Zeiger, P. Barditch-Crovo, and R. F. Siliciano. 1997. Quantification of latent tissue reservoirs and total body viral load in HIV-1 infection. Nature 387:183-188. - PubMed
-
- Chun, T. W., D. Finzi, J. Margolick, K. Chadwick, D. Schwartz, and R. F. Siliciano. 1995. In vivo fate of HIV-1-infected T cells: quantitative analysis of the transition to stable latency. Nat. Med. 1:1284-1290. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
