

mRNA degradation by the virion host shutoff (Vhs) protein of herpes simplex virus: genetic and biochemical evidence that Vhs is a nuclease
- PMID: 12163576
- PMCID: PMC136990
- DOI: 10.1128/jvi.76.17.8560-8571.2002
mRNA degradation by the virion host shutoff (Vhs) protein of herpes simplex virus: genetic and biochemical evidence that Vhs is a nuclease
Abstract
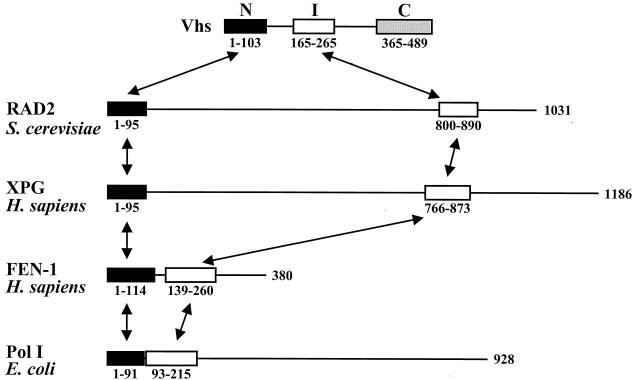
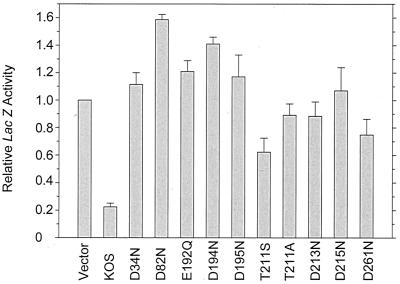
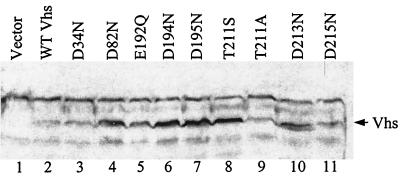
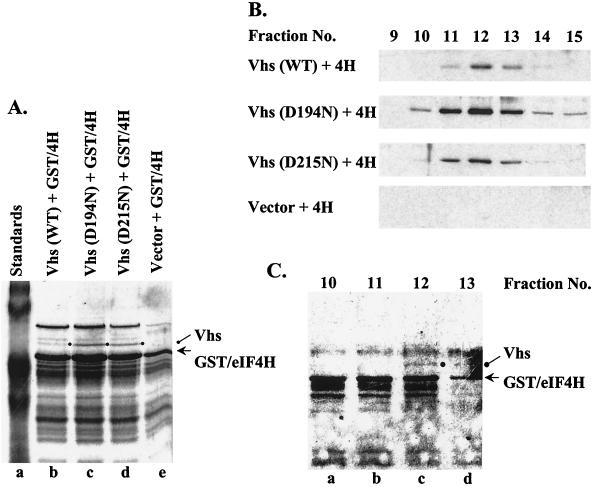
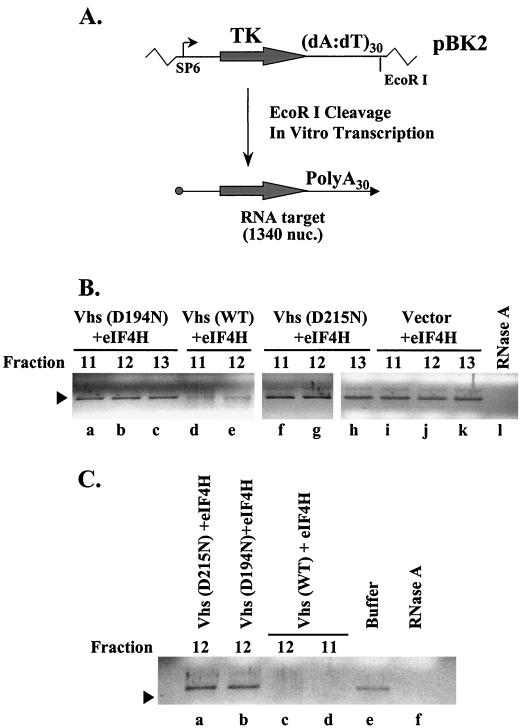
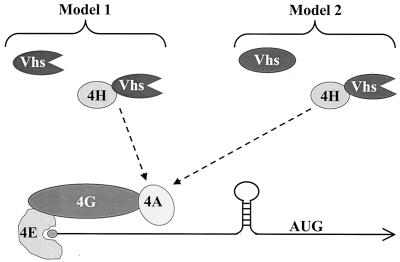
During lytic infections, the virion host shutoff (Vhs) protein (UL41) of herpes simplex virus destabilizes both host and viral mRNAs. By accelerating the decay of all mRNAs, it helps redirect the cell from host to viral gene expression and facilitates the sequential expression of different classes of viral genes. While it is clear that Vhs induces mRNA degradation, it is uncertain whether it is itself an RNase or somehow activates a cellular enzyme. This question was addressed by using a combination of genetic and biochemical approaches. The Vhs homologues of alphaherpesviruses share sequence similarities with a family of mammalian, yeast, bacterial, and phage nucleases. To test the functional significance of these similarities, Vhs was mutated to alter residues corresponding to amino acids known to be critical to the nuclease activity of cellular homologues. In every instance, mutations that inactivated the nuclease activity of cellular homologues also abolished Vhs activity. Recent experiments showed that Vhs interacts with the cellular translation initiation factor eIF4H. In this study, the coexpression of Vhs and a glutathione S-transferase (GST)-eIF4H fusion protein in bacteria resulted in the formation of a complex of the proteins. The wild-type Vhs/GST-eIF4H complex was isolated and shown to have RNase activity. In contrast, Vhs mutations that altered key residues in the nuclease motif abolished the nuclease activity of the recombinant Vhs/GST-eIF4H complex. The results provide genetic and biochemical evidence that Vhs is an RNase, either alone or as a complex with eIF4H.
Figures

References
-
- Becker, Y., E. Tavor, Y. Asher, C. Berkowiltz, and M. Moyal. 1993. Effect of herpes simplex virus type-1 UL41 gene on the stability of mRNA from the cellular genes: beta-actin, fibronectin, glucose transporter-1, and docking protein, and on virus intraperitoneal pathogenicity of newborn mice. Virus Genes 7:133-143. - PubMed
-
- Berthomme, H., B. Jacquemont, and A. Epstein. 1993. The pseudorabies virus host-shutoff homolog gene: nucleotide sequence and comparison with alphaherpesvirus protein counterparts. Virology 193:1028-1032. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
