

Modification of Asn374 of nsP1 suppresses a Sindbis virus nsP4 minus-strand polymerase mutant
- PMID: 12163583
- PMCID: PMC136982
- DOI: 10.1128/jvi.76.17.8641-8649.2002
Modification of Asn374 of nsP1 suppresses a Sindbis virus nsP4 minus-strand polymerase mutant
Abstract
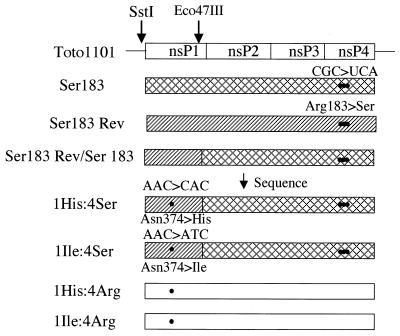
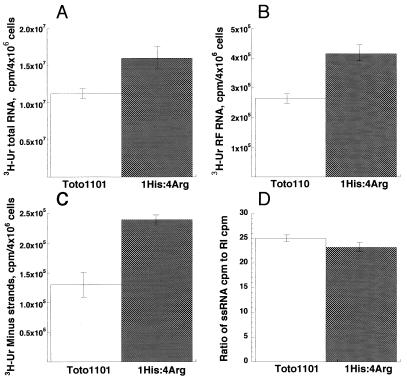
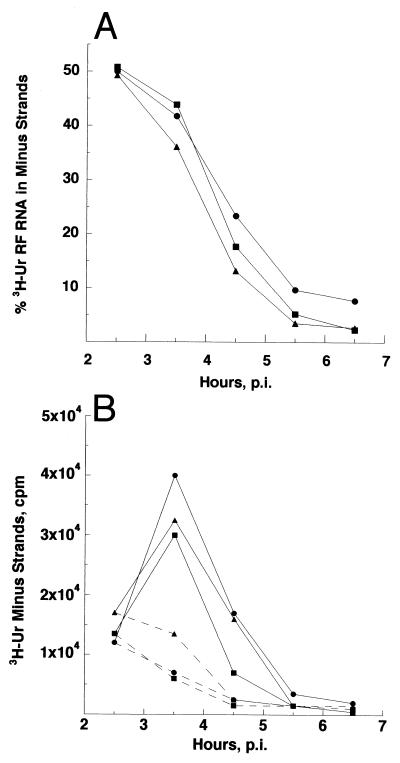
Our recent study (C. L. Fata, S. G. Sawicki, and D. L. Sawicki, J. Virol. 76:8632-8640, 2002) found minus-strand synthesis to be temperature sensitive in vertebrate and invertebrate cells when the Arg183 residue of the Sindbis virus nsP4 polymerase was changed to Ser, Ala, or Lys. Here we report the results of studies identifying an interacting partner of the region of the viral polymerase containing Arg183 that suppresses the Ser183 codon mutation. Large-plaque revertants were observed readily following growth of the nsP4 Ser183 mutant at 40 degrees C. Fifteen revertants were characterized, and all had a mutation in the Asn374 codon of nsP1 that changed it to either a His or an Ile codon. When combined with nsP4 Ser183, substitution of either His374 or Ile374 for Asn374 restored wild-type growth in chicken embryo fibroblast (CEF) cells at 40 degrees C. In Aedes albopictus cells at 34.5 degrees C, neither nsP1 substitution suppressed the nsP4 Ser183 defect in minus-strand synthesis. This argued that the nsP4 Arg183 residue itself is needed for minus-strand replicase assembly or function in the mosquito environment. The nsP1 His374 suppressor when combined with the wild-type nsP4 gave greater than wild-type levels of viral RNA synthesis in CEF cells at 40 degrees C ( approximately 140%) and in Aedes cells at 34.5 degrees C (200%). Virus producing nsP1 His374 and wild-type nsP4 Arg183 made more minus strands during the early period of infection and before minus-strand synthesis ceased at about 4 h postinfection. Shirako et al. (Y. Shirako, E. G. Strauss, and J. H. Strauss, Virology 276:148-160, 2000) identified amino acid substitutions in nsP1 and nsP4 that suppressed mutations that changed the N-terminal Tyr of nsP4. The nsP4 N-terminal mutants were defective also in minus-strand synthesis. Our study implicates an interaction between another conserved nsP1 region and an internal region, predicted to be in the finger domain, of nsP4 for the formation or activity of the minus-strand polymerase. Finally, the observation that a single point mutation in nsP1 results in minus-strand synthesis at greater than wild-type levels supports the concept that the wild-type nsP sequences are evolutionary compromises.
Figures
Similar articles
-
Alphavirus minus-strand RNA synthesis: identification of a role for Arg183 of the nsP4 polymerase.J Virol. 2002 Sep;76(17):8632-40. doi: 10.1128/jvi.76.17.8632-8640.2002. J Virol. 2002. PMID: 12163582 Free PMC article.
-
Suppressor mutations that allow sindbis virus RNA polymerase to function with nonaromatic amino acids at the N-terminus: evidence for interaction between nsP1 and nsP4 in minus-strand RNA synthesis.Virology. 2000 Oct 10;276(1):148-60. doi: 10.1006/viro.2000.0544. Virology. 2000. PMID: 11022003
-
Regulation of Sindbis virus RNA replication: uncleaved P123 and nsP4 function in minus-strand RNA synthesis, whereas cleaved products from P123 are required for efficient plus-strand RNA synthesis.J Virol. 1994 Mar;68(3):1874-85. doi: 10.1128/JVI.68.3.1874-1885.1994. J Virol. 1994. PMID: 8107248 Free PMC article.
-
Alphavirus polymerase and RNA replication.Virus Res. 2017 Apr 15;234:44-57. doi: 10.1016/j.virusres.2017.01.007. Epub 2017 Jan 16. Virus Res. 2017. PMID: 28104453 Review.
-
Alphavirus positive and negative strand RNA synthesis and the role of polyproteins in formation of viral replication complexes.Arch Virol Suppl. 1994;9:393-405. doi: 10.1007/978-3-7091-9326-6_39. Arch Virol Suppl. 1994. PMID: 8032270 Review.
Cited by
-
mRNA capping: biological functions and applications.Nucleic Acids Res. 2016 Sep 19;44(16):7511-26. doi: 10.1093/nar/gkw551. Epub 2016 Jun 17. Nucleic Acids Res. 2016. PMID: 27317694 Free PMC article. Review.
-
Evolutionary genetics and vector adaptation of recombinant viruses of the western equine encephalitis antigenic complex provides new insights into alphavirus diversity and host switching.Virology. 2015 Jan 1;474:154-62. doi: 10.1016/j.virol.2014.10.024. Epub 2014 Nov 17. Virology. 2015. PMID: 25463613 Free PMC article.
-
Inhibition of host protein synthesis by Sindbis virus: correlation with viral RNA replication and release of nuclear proteins to the cytoplasm.Cell Microbiol. 2015 Apr;17(4):520-41. doi: 10.1111/cmi.12381. Epub 2014 Nov 19. Cell Microbiol. 2015. PMID: 25329362 Free PMC article.
-
Increasing the Capping Efficiency of the Sindbis Virus nsP1 Protein Negatively Affects Viral Infection.mBio. 2018 Dec 11;9(6):e02342-18. doi: 10.1128/mBio.02342-18. mBio. 2018. PMID: 30538185 Free PMC article.
-
Role for nsP2 proteins in the cessation of alphavirus minus-strand synthesis by host cells.J Virol. 2006 Jan;80(1):360-71. doi: 10.1128/JVI.80.1.360-371.2006. J Virol. 2006. PMID: 16352561 Free PMC article.
References
-
- Canter, D. M., and J. Perrault. 1996. Stabilization of vesicular stomatitis virus L polymerase protein by P protein binding: a small deletion in the C-terminal domain of L abrogates binding. Virology 219:376-386. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
