

Human immunodeficiency virus type 1 bound to B cells: relationship to virus replicating in CD4+ T cells and circulating in plasma
- PMID: 12163605
- PMCID: PMC136413
- DOI: 10.1128/jvi.76.17.8855-8863.2002
Human immunodeficiency virus type 1 bound to B cells: relationship to virus replicating in CD4+ T cells and circulating in plasma
Abstract
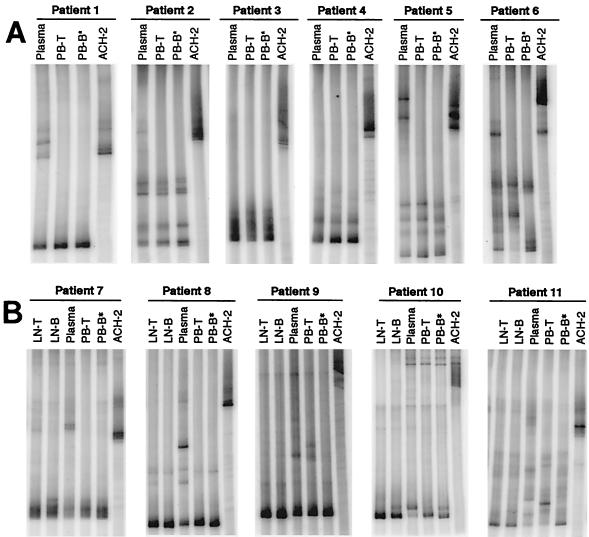
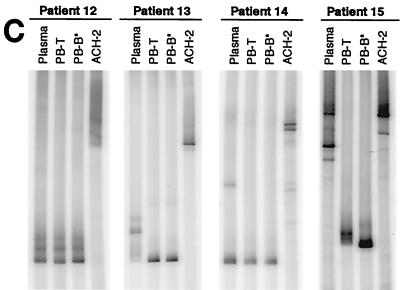
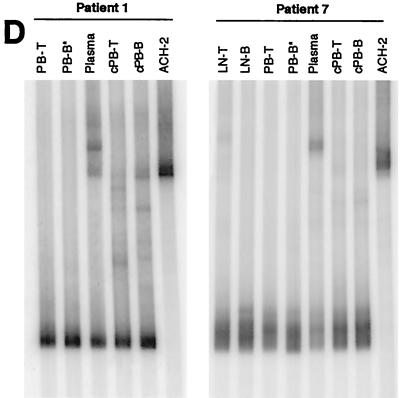
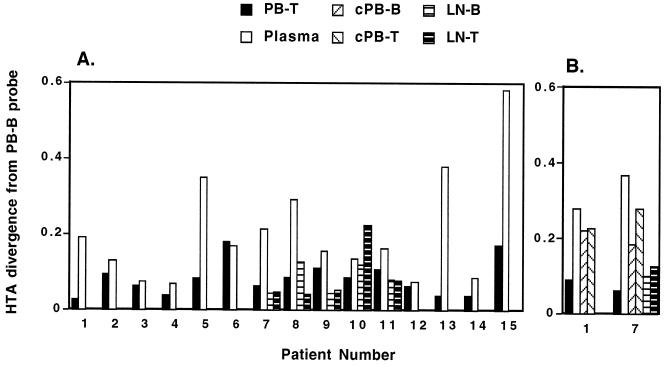
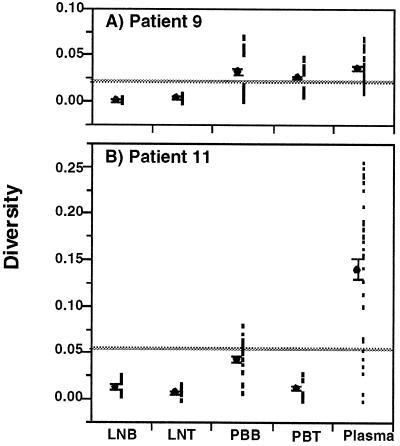
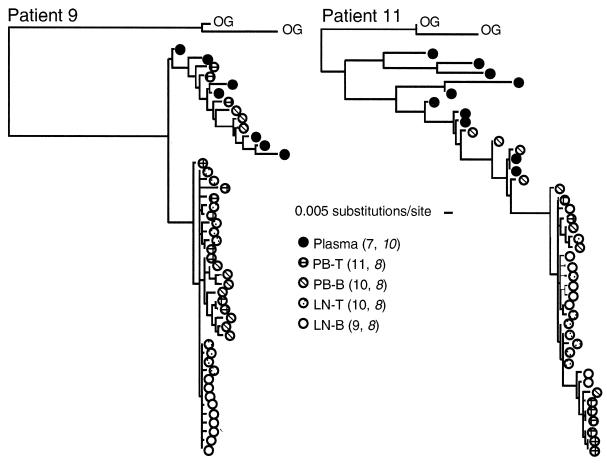
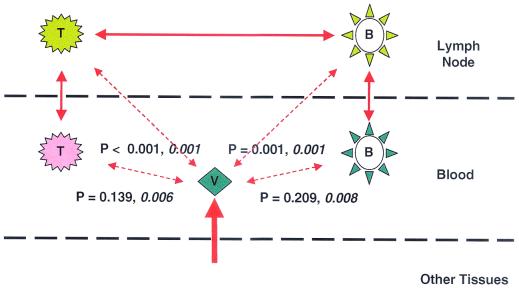
Human immunodeficiency virus type 1 (HIV-1) virions bind to B cells in the peripheral blood and lymph nodes through interactions between CD21 on B cells and complement-complexed virions. B-cell-bound virions have been shown to be highly infectious, suggesting a unique mode of HIV-1 dissemination by B cells circulating between peripheral blood and lymphoid tissues. In order to investigate the relationship between B-cell-bound HIV-1 and viruses found in CD4+ T cells and in plasma, we examined the genetic relationships of HIV-1 found in the blood and lymph nodes of chronically infected patients with heteroduplex mobility and tracking assays and DNA sequence analysis. In samples from 13 of 15 patients examined, HIV-1 variants in peripheral blood-derived B cells were closely related to virus in CD4+ T cells and more divergent from virus in plasma. In samples from five chronically viremic patients for whom analyses were extended to include lymph node-derived HIV-1 isolates, B-cell-associated HIV-1 and CD4+-T-cell-associated HIV-1 in the lymph nodes were equivalent in their divergence from virus in peripheral blood-derived B cells and generally more distantly related to virus in peripheral blood-derived CD4+ T cells. These results indicates virologic cross talk between B cells and CD4+ T cells within the microenvironment of lymphoid tissues and, to a lesser extent, between cells in lymph nodes and the peripheral blood. These findings also indicate that most of the virus in plasma originates from cells other than CD4(+) T cells in the peripheral blood and lymph nodes.
Figures
References
-
- Burton, G. F., A. Masuda, S. L. Heath, B. A. Smith, J. G. Tew, and A. K. Szakal. 1997. Follicular dendritic cells (FDC) in retroviral infection: host/pathogen perspectives. Immunol. Rev. 156:185-197. - PubMed
-
- Chun, T. W., R. T. Davey, Jr., M. Ostrowski, J. Shawn Justement, D. Engel, J. I. Mullins, and A. S. Fauci. 2000. Relationship between pre-existing viral reservoirs and the re-emergence of plasma viremia after discontinuation of highly active anti-retroviral therapy. Nat. Med. 6:757-761. - PubMed
-
- Davey, R. T., Jr., N. Bhat, C. Yoder, T. W. Chun, J. A. Metcalf, R. Dewar, V. Natarajan, R. A. Lempicki, J. W. Adelsberger, K. D. Miller, J. A. Kovacs, M. A. Polis, R. E. Walker, J. Falloon, H. Masur, D. Gee, M. Baseler, D. S. Dimitrov, A. S. Fauci, and H. C. Lane. 1999. HIV-1 and T cell dynamics after interruption of highly active antiretroviral therapy (HAART) in patients with a history of sustained viral suppression. Proc. Natl. Acad. Sci. USA 96:15109-15114. - PMC - PubMed
-
- Delwart, E. L., B. Herring, A. G. Rodrigo, and J. I. Mullins. 1995. Genetic subtyping of human immunodeficiency virus using a heteroduplex mobility assay. PCR Methods Appl. 4:S202-S216. - PubMed
-
- Delwart, E. L., and C. J. Gordon. 1997. Tracking changes in HIV-1 envelope quasispecies using DNA heteroduplex analysis. Methods 12:348-354. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
