

Inactivation of NF-kappaB-dependent cell survival, a novel mechanism for the proapoptotic function of c-Abl
- PMID: 12167702
- PMCID: PMC134000
- DOI: 10.1128/MCB.22.17.6079-6088.2002
Inactivation of NF-kappaB-dependent cell survival, a novel mechanism for the proapoptotic function of c-Abl
Abstract
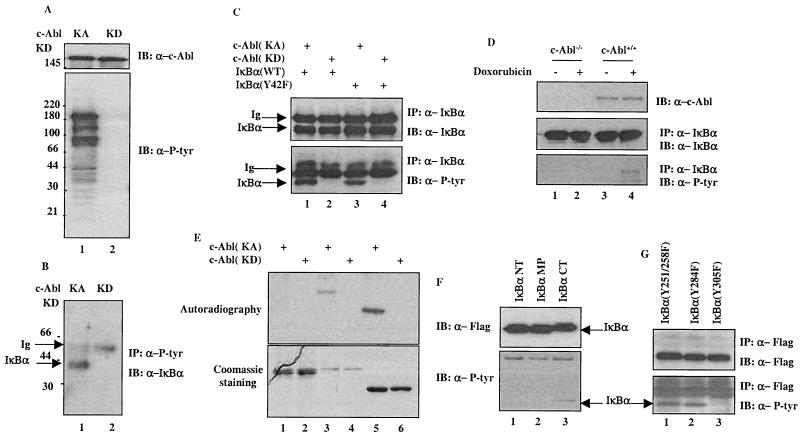
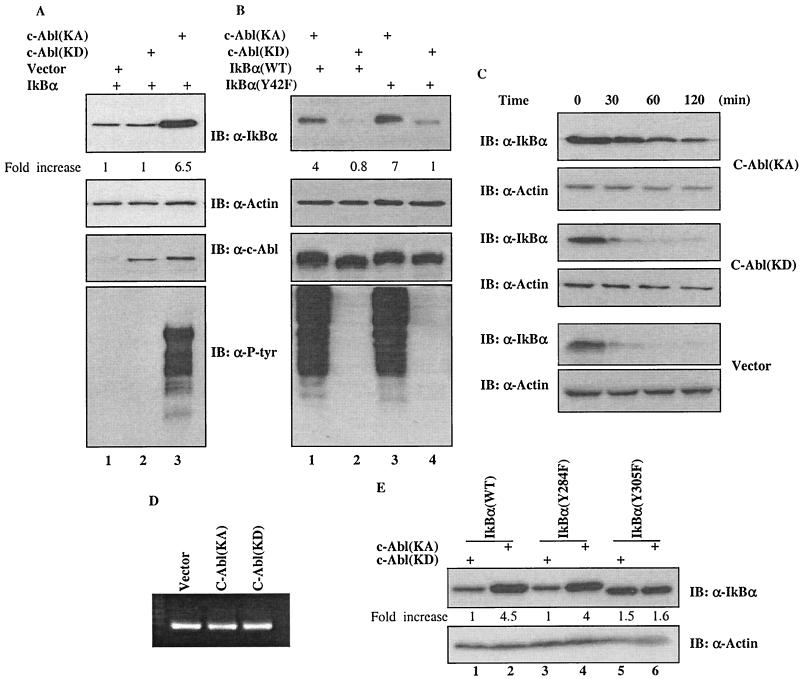
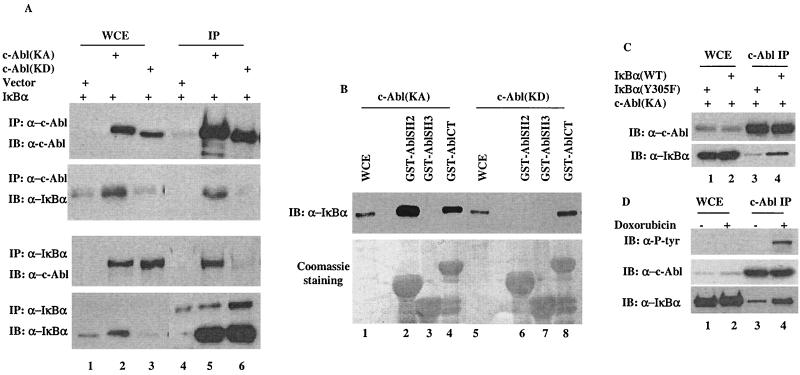
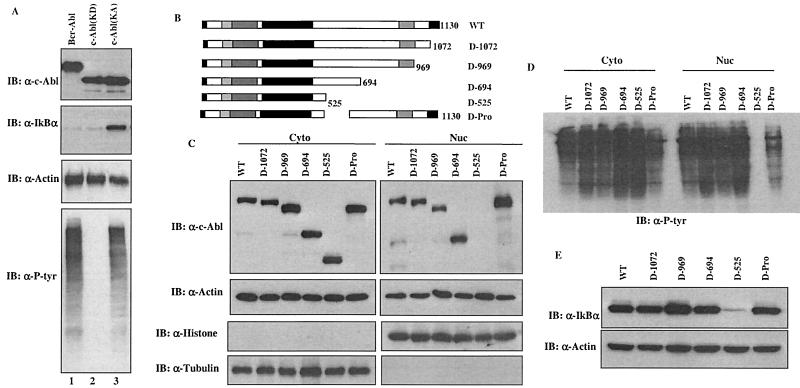
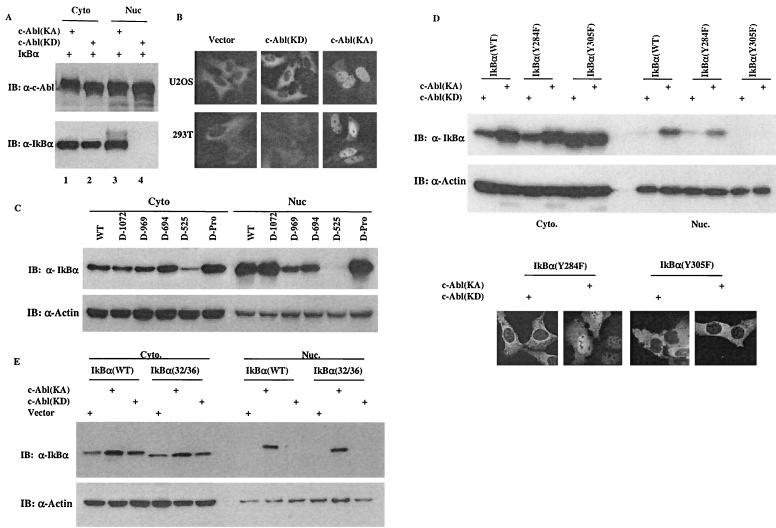
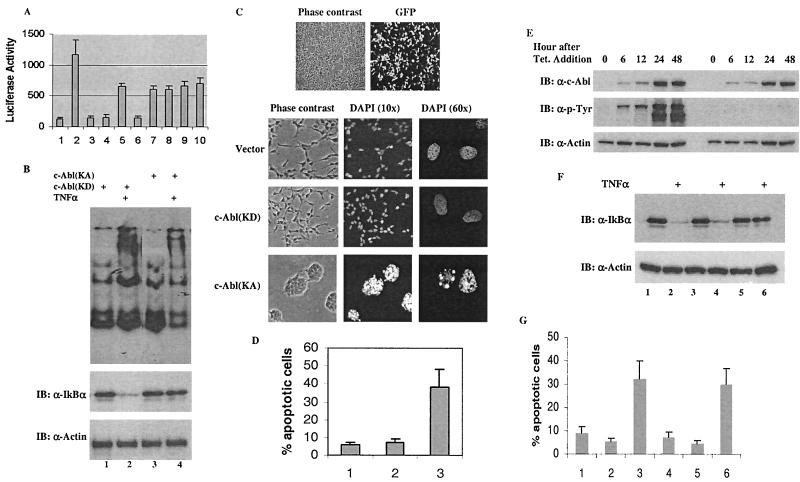
Using a system that expresses a constitutively kinase-active c-Abl protein [c-Abl(KA)], we identified the protein IkappaBalpha as a novel substrate of c-Abl. This kinase-substrate relationship is not only confirmed at the level of endogenous proteins but is also supported by a physical interaction between the two proteins. Interestingly, the association of c-Abl with IkappaBalpha, which is detectable in the form of nonphosphorylated proteins, is remarkably enhanced by an inducible binding of tyrosine-phosphorylated IkappaBalpha to the c-Abl SH2 domain. In contrast to the serine 32/34 phosphorylation that triggers ubiquitination and degradation of IkappaBalpha, c-Abl-mediated phosphorylation at tyrosine 305 is associated with an increase of the IkappaBalpha protein stability. Significantly, this activity is not shared by the oncogenic Bcr-Abl, because it is unique to the nuclear c-Abl. We also demonstrate that c-Abl targets the nuclear subpopulation of IkappaBalpha for phosphorylation and induces it to accumulate in the nucleus. As a consequence, NF-kappaB transcription activity is abolished, leading to an increased cellular sensitivity to the induction of apoptosis. The functional importance of c-Abl-mediated IkappaBalpha phosphorylation is highlighted by a loss of response of the IkappaBalpha(Y305F) protein to c-Abl-mediated regulation. Using cells expressing the c-Abl(KA) protein under the control of an inducible promoter, we demonstrate inactivation of the NF-kappaB-dependent cell survival pathway as one of the mechanisms for c-Abl-mediated apoptosis.
Figures
Similar articles
-
Ectopic expression of protein-tyrosine kinase Bcr-Abl suppresses tumor necrosis factor (TNF)-induced NF-kappa B activation and IkappaBalpha phosphorylation. Relationship with down-regulation of TNF receptors.J Biol Chem. 2002 Aug 23;277(34):30622-8. doi: 10.1074/jbc.M204748200. Epub 2002 Jun 11. J Biol Chem. 2002. PMID: 12060665
-
Doxorubicin induces atypical NF-κB activation through c-Abl kinase activity in breast cancer cells.J Cancer Res Clin Oncol. 2013 Oct;139(10):1625-35. doi: 10.1007/s00432-013-1476-3. Epub 2013 Jul 28. J Cancer Res Clin Oncol. 2013. PMID: 23892407 Free PMC article.
-
Regulation of p73 by c-Abl through the p38 MAP kinase pathway.Oncogene. 2002 Jan 31;21(6):974-9. doi: 10.1038/sj.onc.1205134. Oncogene. 2002. PMID: 11840343
-
Molecular biology of chronic myeloid leukemia.Int J Hematol. 2001 Apr;73(3):308-22. doi: 10.1007/BF02981955. Int J Hematol. 2001. PMID: 11345196 Review.
-
ABL tyrosine kinases: evolution of function, regulation, and specificity.Sci Signal. 2010 Sep 14;3(139):re6. doi: 10.1126/scisignal.3139re6. Sci Signal. 2010. PMID: 20841568 Free PMC article. Review.
Cited by
-
Activation of NF-κB in B cell receptor signaling through Bruton's tyrosine kinase-dependent phosphorylation of IκB-α.J Mol Med (Berl). 2019 May;97(5):675-690. doi: 10.1007/s00109-019-01777-x. Epub 2019 Mar 19. J Mol Med (Berl). 2019. PMID: 30887112
-
Redox-regulated pathway of tyrosine phosphorylation underlies NF-κB induction by an atypical pathway independent of the 26S proteasome.Biomolecules. 2015 Feb 9;5(1):95-112. doi: 10.3390/biom5010095. Biomolecules. 2015. PMID: 25671697 Free PMC article.
-
Genotoxicity-Stimulated and CYLD-Driven Malignant Transformation.Cancer Manag Res. 2022 Aug 5;14:2339-2356. doi: 10.2147/CMAR.S373557. eCollection 2022. Cancer Manag Res. 2022. PMID: 35958947 Free PMC article.
-
Abelson virus transformation prevents TRAIL expression by inhibiting FoxO3a and NF-kappaB.Mol Cells. 2010 Apr;29(4):333-41. doi: 10.1007/s10059-010-0029-8. Epub 2010 Mar 4. Mol Cells. 2010. PMID: 20213318 Free PMC article.
-
Growth inhibitory effect of dihydroartemisinin on Bcr/Abl+ chronic myeloid leukemia K562 cells involve AKT, ERK and NF-κB modulation.J Cancer Res Clin Oncol. 2012 Dec;138(12):2095-102. doi: 10.1007/s00432-012-1292-1. Epub 2012 Jul 26. J Cancer Res Clin Oncol. 2012. PMID: 22833150 Free PMC article.
References
-
- Aradhya, S., and D. L. Nelson. 2000. NF-κB signaling and human disease. Curr. Opin. Genet. Dev. 11:300-306. - PubMed
-
- Costanzo, A., P. Merlo, N. Pediconi, M. Fulco, V. Sartorelli, P. A. Cole, G. Fontemaggi, M. Fanciulli, L. Schiltz, G. Blandino, C. Balsano, and M. Levrero. 2002. DNA damage-dependent acetylation of p73 dictates the selective activation of apoptotic target genes. Mol. Cell 9:175-186. - PubMed
-
- Huxford, T., S. Malek, and G. Ghosh. 1999. Structure and mechanism in NF-kappa B/I kappa B signaling. Cold Spring Harbor Symp. Quant. Biol. 64: 533-540. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous