

Phospholemman (FXYD1) associates with Na,K-ATPase and regulates its transport properties
- PMID: 12169672
- PMCID: PMC123281
- DOI: 10.1073/pnas.182267299
Phospholemman (FXYD1) associates with Na,K-ATPase and regulates its transport properties
Abstract
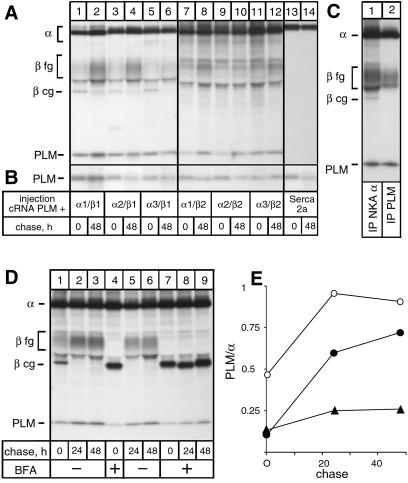
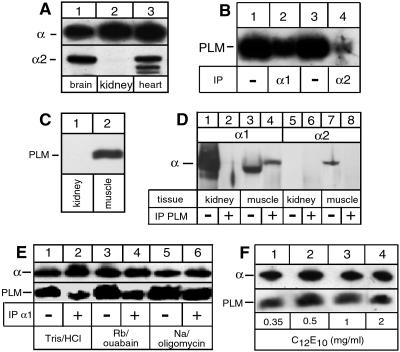
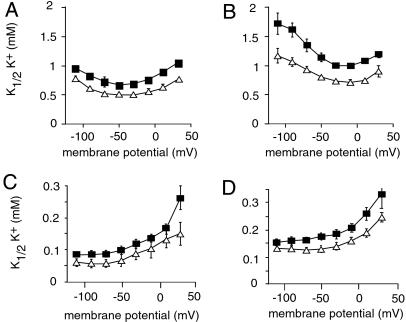
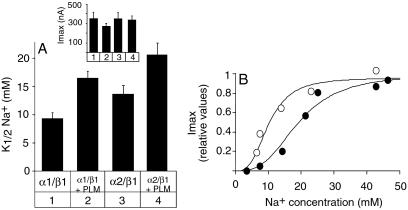
A family of small, single-span membrane proteins (the FXYD family) has recently been defined based on their sequence and structural homology. Some members of this family have already been identified as tissue-specific regulators of Na,K-ATPase (NKA). In the present study, we demonstrate that phospholemman (PLM) (FXYD1), so far considered to be a heart- and muscle-specific channel or channel-regulating protein, associates specifically and stably with six different alpha-beta isozymes of NKA after coexpression in Xenopus oocytes, and with alpha1-beta, and less efficiently with alpha2-beta isozymes, in native cardiac and skeletal muscles. Stoichiometric association of PLM with NKA occurs posttranslationally either in the Golgi or the plasma membrane. Interaction of PLM with NKA induces a small decrease in the external K+ affinity of alpha1-beta1 and alpha2-beta1 isozymes and a nearly 2-fold decrease in the internal Na+ affinity. In conclusion, this study demonstrates that PLM is a tissue-specific regulator of NKA that may play an essential role in muscle contractility.
Figures
Similar articles
-
Phosphorylation of phospholemman (FXYD1) by protein kinases A and C modulates distinct Na,K-ATPase isozymes.J Biol Chem. 2008 Jan 4;283(1):476-486. doi: 10.1074/jbc.M705830200. Epub 2007 Nov 8. J Biol Chem. 2008. PMID: 17991751
-
Intracellular trafficking of FXYD1 (phospholemman) and FXYD7 proteins in Xenopus oocytes and mammalian cells.J Biol Chem. 2012 Jun 15;287(25):21130-41. doi: 10.1074/jbc.M112.347807. Epub 2012 Apr 25. J Biol Chem. 2012. PMID: 22535957 Free PMC article.
-
Isoform specificity of the Na/K-ATPase association and regulation by phospholemman.J Biol Chem. 2009 Sep 25;284(39):26749-57. doi: 10.1074/jbc.M109.047357. Epub 2009 Jul 28. J Biol Chem. 2009. PMID: 19638348 Free PMC article.
-
Coordinated regulation of cardiac Na(+)/Ca (2+) exchanger and Na (+)-K (+)-ATPase by phospholemman (FXYD1).Adv Exp Med Biol. 2013;961:175-90. doi: 10.1007/978-1-4614-4756-6_15. Adv Exp Med Biol. 2013. PMID: 23224879 Review.
-
FXYD proteins: new tissue- and isoform-specific regulators of Na,K-ATPase.Ann N Y Acad Sci. 2003 Apr;986:388-94. doi: 10.1111/j.1749-6632.2003.tb07219.x. Ann N Y Acad Sci. 2003. PMID: 12763855 Review.
Cited by
-
Pivotal role of α2 Na+ pumps and their high affinity ouabain binding site in cardiovascular health and disease.J Physiol. 2016 Nov 1;594(21):6079-6103. doi: 10.1113/JP272419. Epub 2016 Jul 31. J Physiol. 2016. PMID: 27350568 Free PMC article. Review.
-
Different Modulatory Mechanisms of Renal FXYD12 for Na(+)-K(+)-ATPase between Two Closely Related Medakas upon Salinity Challenge.Int J Biol Sci. 2016 Apr 28;12(6):730-45. doi: 10.7150/ijbs.15066. eCollection 2016. Int J Biol Sci. 2016. PMID: 27194950 Free PMC article.
-
Muscle variables of importance for physiological performance in competitive football.Eur J Appl Physiol. 2016 Feb;116(2):251-62. doi: 10.1007/s00421-015-3274-x. Epub 2015 Oct 8. Eur J Appl Physiol. 2016. PMID: 26449984
-
Data-Independent Acquisition Proteomics and N-Terminomics Methods Reveal Alterations in Mitochondrial Function and Metabolism in Ischemic-Reperfused Hearts.J Proteome Res. 2024 Feb 2;23(2):844-856. doi: 10.1021/acs.jproteome.3c00754. Epub 2024 Jan 24. J Proteome Res. 2024. PMID: 38264990 Free PMC article.
-
Regulation of the cardiac sodium pump.Cell Mol Life Sci. 2013 Apr;70(8):1357-80. doi: 10.1007/s00018-012-1134-y. Epub 2012 Sep 7. Cell Mol Life Sci. 2013. PMID: 22955490 Free PMC article. Review.
References
-
- Gheorghiade M. & Pitt, B. (1997) Am. Heart J. 134, 3-12. - PubMed
-
- Hamlyn J. M., Hamilton, B. P. & Manunta, P. (1996) J. Hypertens. 14, 151-167. - PubMed
-
- Geering K. (2001) J. Bioenerg. Biomembr. 33, 425-438. - PubMed
-
- Crambert G., Hasler, U., Beggah, A. T., Yu, C., Modyanov, N. N., Horisberger, J. D., Lelièvre, L. & Geering, K. (2000) J. Biol. Chem. 275, 1976-1986. - PubMed
-
- Blanco G. & Mercer, R. W. (1998) Am. J. Physiol. 275, F633-F650. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
