

A major light-harvesting polypeptide of photosystem II functions in thermal dissipation
- PMID: 12172023
- PMCID: PMC151466
- DOI: 10.1105/tpc.002154
A major light-harvesting polypeptide of photosystem II functions in thermal dissipation
Abstract
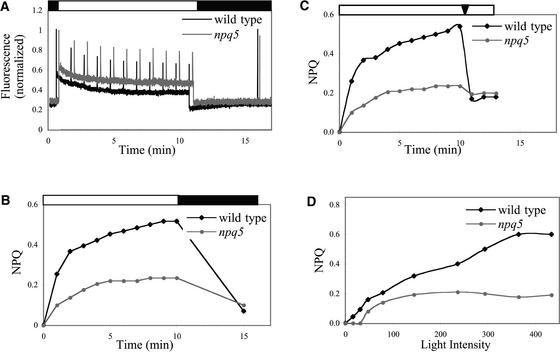
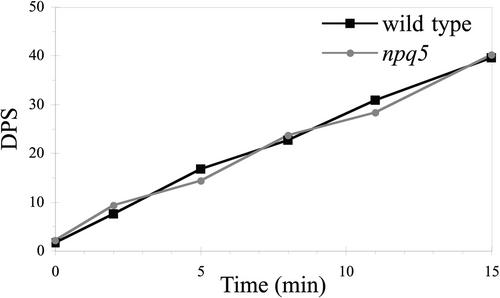
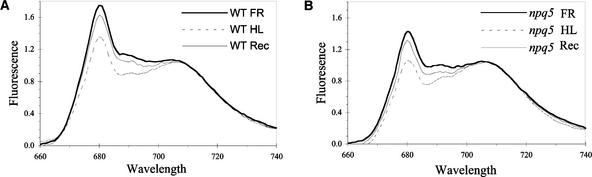
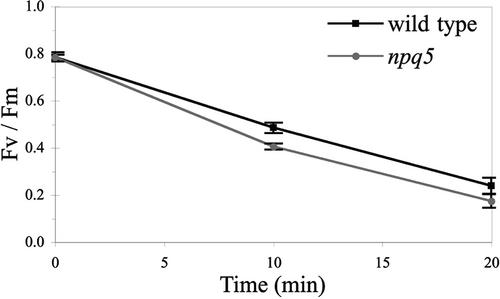
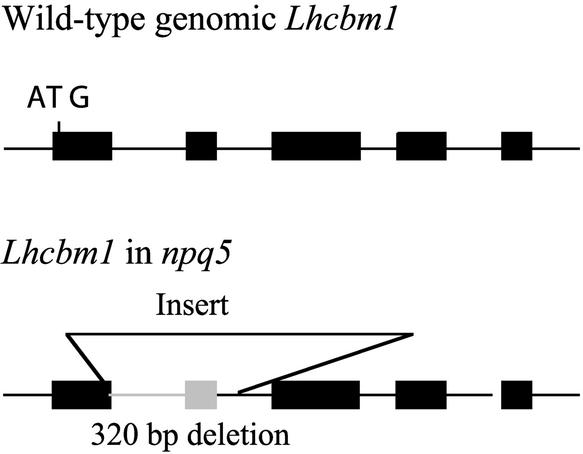
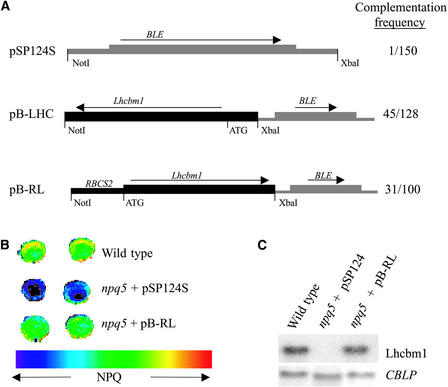
Under high-light conditions, photoprotective mechanisms minimize the damaging effects of excess light. A primary photoprotective mechanism is thermal dissipation of excess excitation energy within the light-harvesting complex of photosystem II (LHCII). Although roles for both carotenoids and specific polypeptides in thermal dissipation have been reported, neither the site nor the mechanism of this process has been defined precisely. Here, we describe the physiological and molecular characteristics of the Chlamydomonas reinhardtii npq5 mutant, a strain that exhibits little thermal dissipation. This strain is normal for state transition, high light-induced violaxanthin deepoxidation, and low light growth, but it is more sensitive to photoinhibition than the wild type. Furthermore, both pigment data and measurements of photosynthesis suggest that the photosystem II antenna in the npq5 mutant has one-third fewer light-harvesting trimers than do wild-type cells. The npq5 mutant is null for a gene designated Lhcbm1, which encodes a light-harvesting polypeptide present in the trimers of the photosystem II antennae. Based on sequence data, the Lhcbm1 gene is 1 of 10 genes that encode the major LHCII polypeptides in Chlamydomonas. Amino acid alignments demonstrate that these predicted polypeptides display a high degree of sequence identity but maintain specific differences in their N-terminal regions. Both physiological and molecular characterization of the npq5 mutant suggest that most thermal dissipation within LHCII of Chlamydomonas is dependent on the peripherally associated trimeric LHC polypeptides.
Figures

Comment in
-
A role for a light-harvesting antenna complex of photosystem II in photoprotection.Plant Cell. 2002 Aug;14(8):1663-8. doi: 10.1105/tpc.140810. Plant Cell. 2002. PMID: 12172013 Free PMC article. No abstract available.
Similar articles
-
Photosynthesis, chlorophyll fluorescence, light-harvesting system and photoinhibition resistance of a zeaxanthin-accumulating mutant of Arabidopsis thaliana.J Photochem Photobiol B. 1996 Jun;34(1):87-94. doi: 10.1016/1011-1344(95)07272-1. J Photochem Photobiol B. 1996. PMID: 8765663
-
Absence of lutein, violaxanthin and neoxanthin affects the functional chlorophyll antenna size of photosystem-II but not that of photosystem-I in the green alga Chlamydomonas reinhardtii.Plant Cell Physiol. 2001 May;42(5):482-91. doi: 10.1093/pcp/pce058. Plant Cell Physiol. 2001. PMID: 11382814
-
A role for a light-harvesting antenna complex of photosystem II in photoprotection.Plant Cell. 2002 Aug;14(8):1663-8. doi: 10.1105/tpc.140810. Plant Cell. 2002. PMID: 12172013 Free PMC article. No abstract available.
-
A genome's-eye view of the light-harvesting polypeptides of Chlamydomonas reinhardtii.Curr Genet. 2004 Feb;45(2):61-75. doi: 10.1007/s00294-003-0460-x. Epub 2003 Dec 2. Curr Genet. 2004. PMID: 14652691 Review.
-
Molecular design of the photosystem II light-harvesting antenna: photosynthesis and photoprotection.J Exp Bot. 2005 Jan;56(411):365-73. doi: 10.1093/jxb/eri023. Epub 2004 Nov 22. J Exp Bot. 2005. PMID: 15557295 Review.
Cited by
-
Correlation between spatial (3D) structure of pea and bean thylakoid membranes and arrangement of chlorophyll-protein complexes.BMC Plant Biol. 2012 May 25;12:72. doi: 10.1186/1471-2229-12-72. BMC Plant Biol. 2012. PMID: 22631450 Free PMC article.
-
A mechanism of nonphotochemical energy dissipation, independent from PsbS, revealed by a conformational change in the antenna protein CP26.Plant Cell. 2005 Apr;17(4):1217-32. doi: 10.1105/tpc.104.030601. Epub 2005 Mar 4. Plant Cell. 2005. PMID: 15749754 Free PMC article.
-
Interactions between the photosystem II subunit PsbS and xanthophylls studied in vivo and in vitro.J Biol Chem. 2008 Mar 28;283(13):8434-45. doi: 10.1074/jbc.M708291200. Epub 2007 Dec 10. J Biol Chem. 2008. PMID: 18070876 Free PMC article.
-
Hydrogen photoproduction is attenuated by disruption of an isoamylase gene in Chlamydomonas reinhardtii.Plant Cell. 2004 Aug;16(8):2151-63. doi: 10.1105/tpc.104.021972. Epub 2004 Jul 21. Plant Cell. 2004. PMID: 15269330 Free PMC article.
-
Analysis of LhcSR3, a protein essential for feedback de-excitation in the green alga Chlamydomonas reinhardtii.PLoS Biol. 2011 Jan 18;9(1):e1000577. doi: 10.1371/journal.pbio.1000577. PLoS Biol. 2011. PMID: 21267060 Free PMC article.
References
-
- Andrews, J.R., Fryer, M.J., and Baker, N.R. (1995). Consequences of LHCII deficiency for photosynthetic regulation in chlorina mutants of barley. Photosynth. Res. 44, 81–91. - PubMed
-
- Asada, K. (1999). The water-water cycle in chloroplasts: Scavenging of active oxygen and dissipation of excess photons. Annu. Rev. Plant Physiol. Plant Mol. Biol. 50, 601–639. - PubMed
-
- Bassi, R., Giacometti, G.M., and Simpson, D.J. (1988). Changes in the organization of stroma membranes induced by in vivo state- 1-state-2 transition. Biochim. Biophys. Acta 935, 152–165.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
