

VGF is required for obesity induced by diet, gold thioglucose treatment, and agouti and is differentially regulated in pro-opiomelanocortin- and neuropeptide Y-containing arcuate neurons in response to fasting
- PMID: 12177191
- PMCID: PMC6757869
- DOI: 10.1523/JNEUROSCI.22-16-06929.2002
VGF is required for obesity induced by diet, gold thioglucose treatment, and agouti and is differentially regulated in pro-opiomelanocortin- and neuropeptide Y-containing arcuate neurons in response to fasting
Abstract
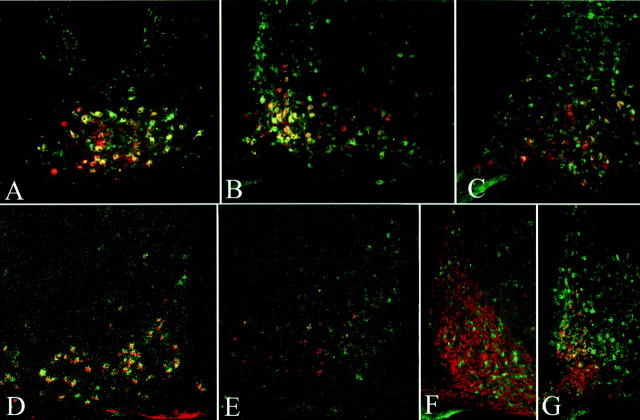
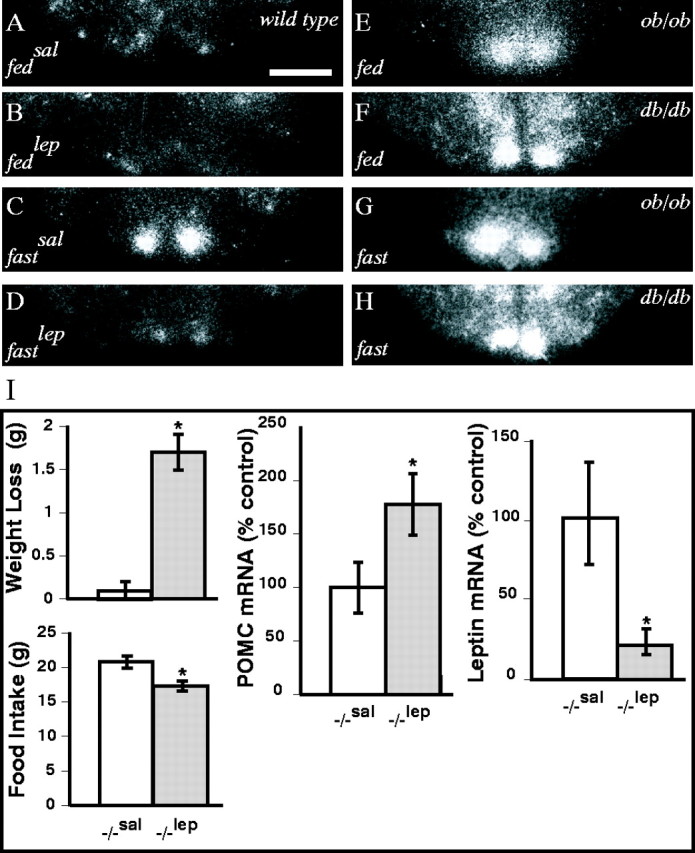
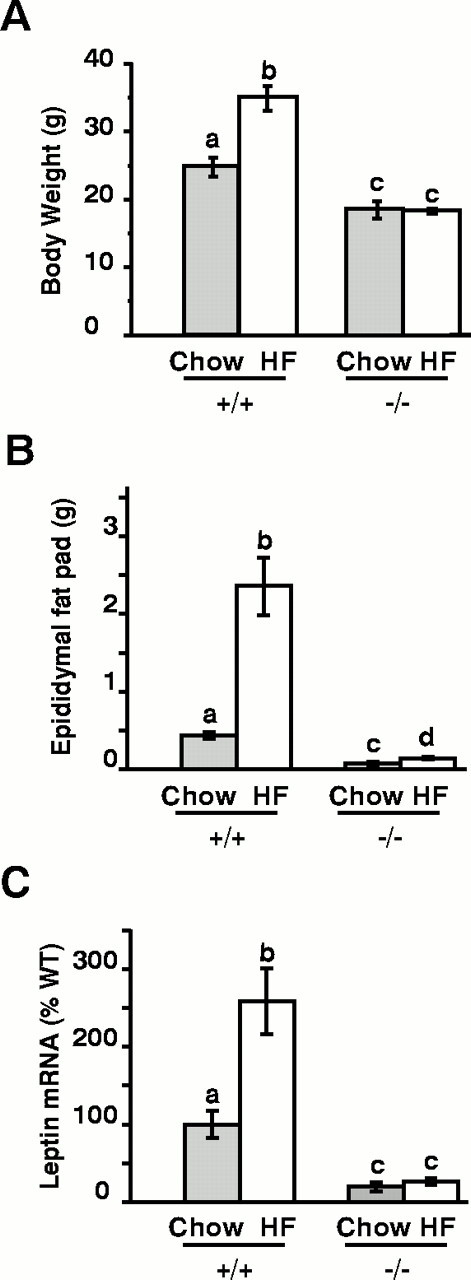
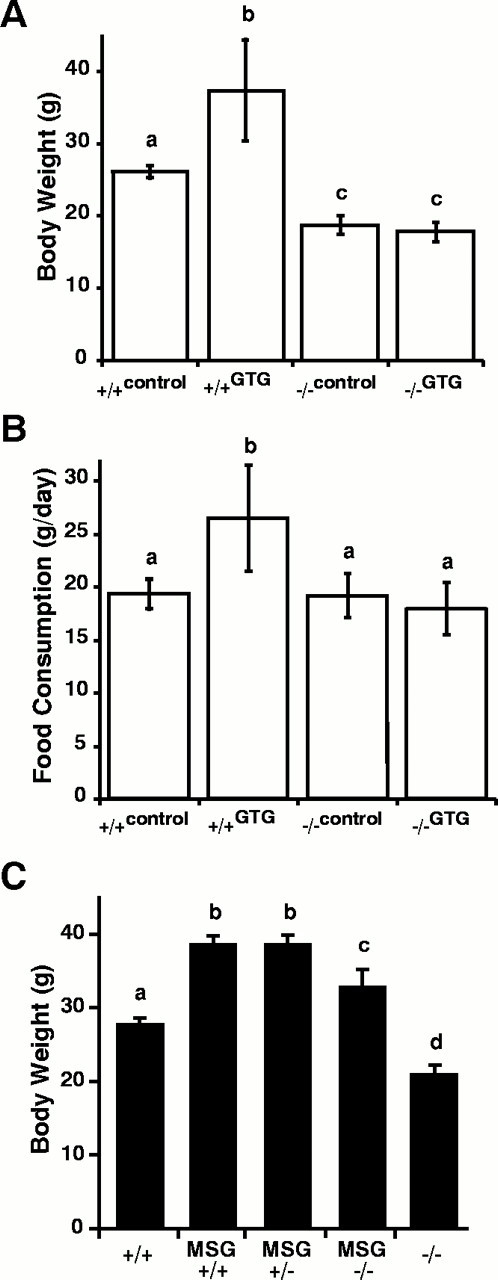
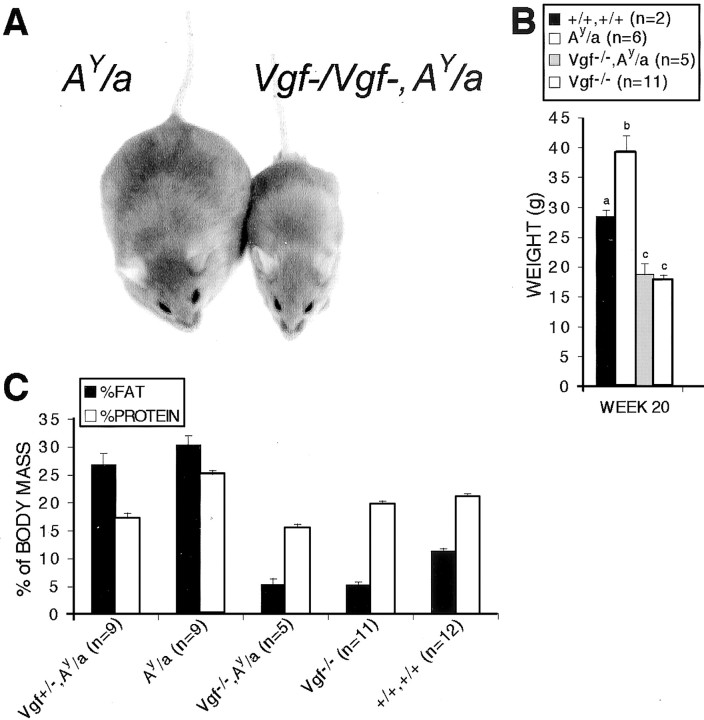
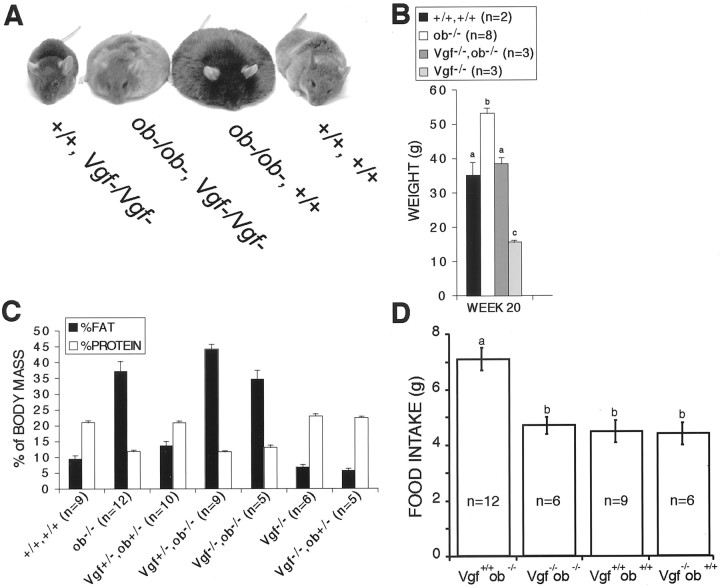
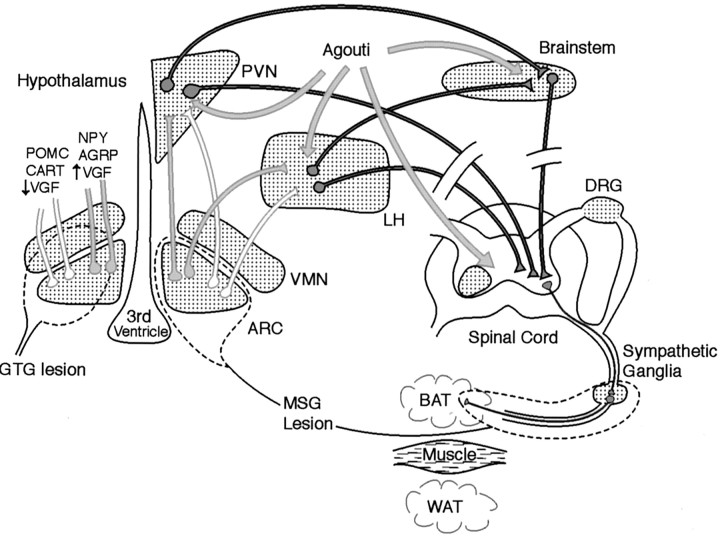
Targeted deletion of the gene encoding the neuronal and neuroendocrine secreted polypeptide VGF (nonacronymic) produces a lean, hypermetabolic mouse. Consistent with this phenotype, VGF mRNA levels are regulated in the hypothalamic arcuate nucleus in response to fasting. To gain insight into the site(s) and mechanism(s) of action of VGF, we further characterized VGF expression in the hypothalamus. Double-label studies indicated that VGF and pro-opiomelanocortin were coexpressed in lateral arcuate neurons in the fed state, and that VGF expression was induced after fasting in medial arcuate neurons that synthesize neuropeptide Y (NPY). Like NPY, VGF mRNA induction in this region of the hypothalamus in fasted mice was inhibited by exogenous leptin. In leptin-deficient ob/ob and receptor-mutant db/db mice, VGF mRNA levels in the medial arcuate were elevated. To identify neural pathways that are functionally compromised by Vgf ablation, VGF mutant mice were crossed with obese A(y)/a (agouti) and ob/ob mice. VGF deficiency completely blocked the development of obesity in A(y)/a mice, whereas deletion of Vgf in ob/ob mice attenuated weight gain but had no impact on adiposity. Hypothalamic levels of NPY and agouti-related polypeptide mRNAs in both double-mutant lines were dramatically elevated 10- to 15-fold above those of wild-type mice. VGF-deficient mice were also found to resist diet- and gold thioglucose-induced obesity. These data and the susceptibility of VGF mutant mice to monosodium glutamate-induced obesity are consistent with a role for VGF in outflow pathways, downstream of hypothalamic and/or brainstem melanocortin 4 receptors, that project via the autonomic nervous system to peripheral metabolic tissues and regulate energy homeostasis.
Figures
Similar articles
-
Targeted deletion of the Vgf gene indicates that the encoded secretory peptide precursor plays a novel role in the regulation of energy balance.Neuron. 1999 Jul;23(3):537-48. doi: 10.1016/s0896-6273(00)80806-5. Neuron. 1999. PMID: 10433265
-
Leptin increases hypothalamic pro-opiomelanocortin mRNA expression in the rostral arcuate nucleus.Diabetes. 1997 Dec;46(12):2119-23. doi: 10.2337/diab.46.12.2119. Diabetes. 1997. PMID: 9392508
-
Induction of neuropeptide Y gene expression in the dorsal medial hypothalamic nucleus in two models of the agouti obesity syndrome.Mol Endocrinol. 1997 May;11(5):630-7. doi: 10.1210/mend.11.5.9921. Mol Endocrinol. 1997. PMID: 9139806
-
VGF: a novel role for this neuronal and neuroendocrine polypeptide in the regulation of energy balance.Front Neuroendocrinol. 2000 Jul;21(3):199-219. doi: 10.1006/frne.2000.0199. Front Neuroendocrinol. 2000. PMID: 10882540 Review.
-
The hypothalamus and the regulation of energy homeostasis: lifting the lid on a black box.Proc Nutr Soc. 2000 Aug;59(3):385-96. doi: 10.1017/s0029665100000434. Proc Nutr Soc. 2000. PMID: 10997654 Review.
Cited by
-
Arctic charr brain transcriptome strongly affected by summer seasonal growth but only subtly by feed deprivation.BMC Genomics. 2019 Jun 27;20(1):529. doi: 10.1186/s12864-019-5874-z. BMC Genomics. 2019. PMID: 31248377 Free PMC article.
-
The granin VGF promotes genesis of secretory vesicles, and regulates circulating catecholamine levels and blood pressure.FASEB J. 2014 May;28(5):2120-33. doi: 10.1096/fj.13-239509. Epub 2014 Feb 4. FASEB J. 2014. PMID: 24497580 Free PMC article.
-
Processing, distribution, and function of VGF, a neuronal and endocrine peptide precursor.Cell Mol Neurobiol. 2004 Aug;24(4):517-33. doi: 10.1023/b:cemn.0000023627.79947.22. Cell Mol Neurobiol. 2004. PMID: 15233376 Free PMC article. Review.
-
Germline ablation of VGF increases lipolysis in white adipose tissue.J Endocrinol. 2012 Nov;215(2):313-22. doi: 10.1530/JOE-12-0172. Epub 2012 Aug 31. J Endocrinol. 2012. PMID: 22942234 Free PMC article.
-
Age-related increase of VGF-expression in T lymphocytes.Aging (Albany NY). 2014 Jun;6(6):440-53. doi: 10.18632/aging.100656. Aging (Albany NY). 2014. PMID: 25013207 Free PMC article.
References
-
- Ahima RS, Saper CB, Flier JS, Elmquist JK. Leptin regulation of neuroendocrine systems. Front Neuroendocrinol. 2000;21:263–307. - PubMed
-
- Bamshad M, Song CK, Bartness TJ. CNS origins of the sympathetic nervous system outflow to brown adipose tissue. Am J Physiol. 1999;276:R1569–R1578. - PubMed
-
- Barsh G. From Agouti to Pomc: 100 years of fat blonde mice. Nat Med. 1999;5:984–985. - PubMed
-
- Barsh GS, Farooqi IS, O'Rahilly S. Genetics of body-weight regulation. Nature. 2000;404:644–651. - PubMed
-
- Bergen HT, Mizuno TM, Taylor J, Mobbs CV. Hyperphagia and weight gain after gold-thioglucose: relation to hypothalamic neuropeptide Y and proopiomelanocortin. Endocrinology. 1998;139:4483–4488. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous