

Caspase 3 activity is required for skeletal muscle differentiation
- PMID: 12177420
- PMCID: PMC123204
- DOI: 10.1073/pnas.162172899
Caspase 3 activity is required for skeletal muscle differentiation
Abstract
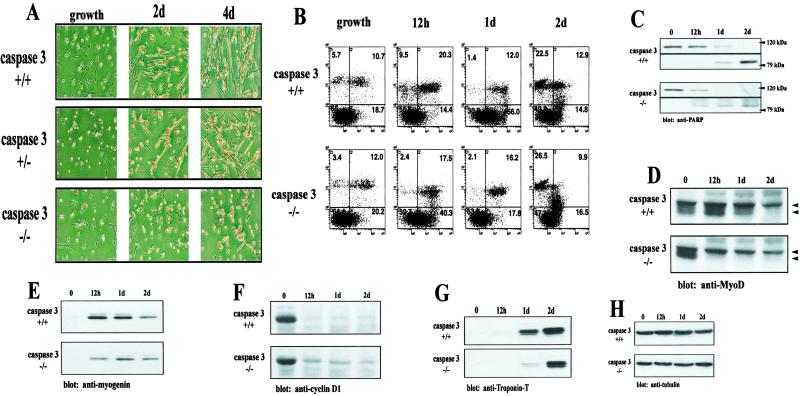
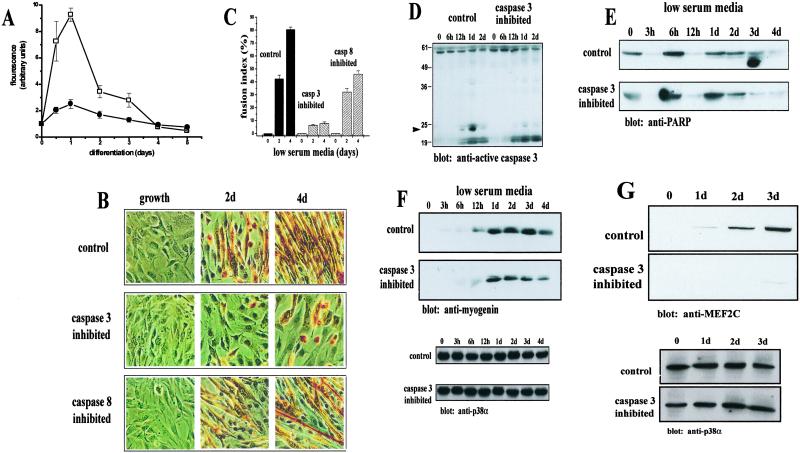
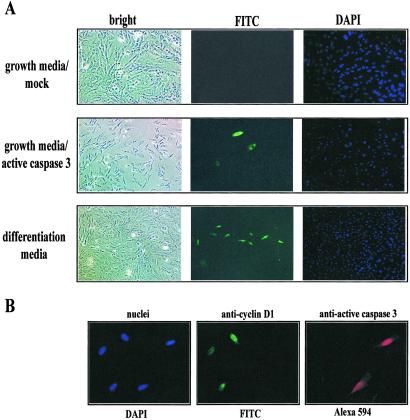
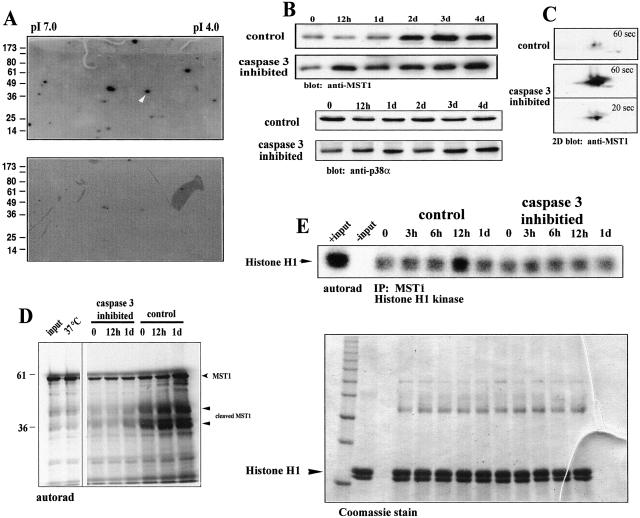
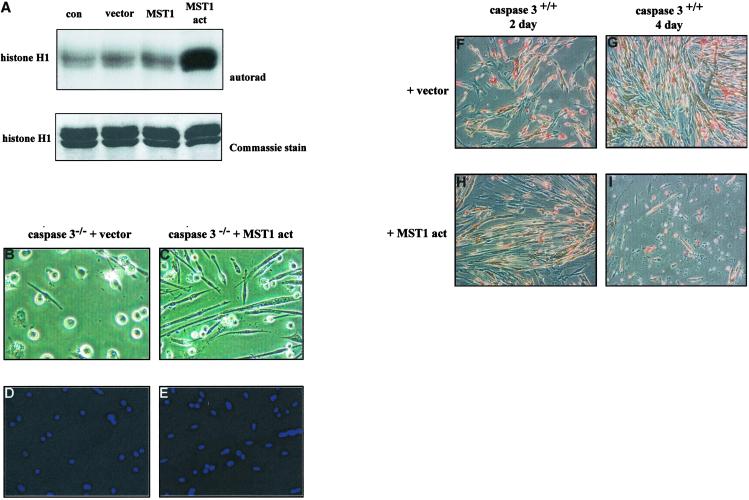
The cellular alterations associated with skeletal muscle differentiation share a high degree of similarity with key phenotypic changes usually ascribed to apoptosis. For example, actin fiber disassembly/reorganization is a conserved feature of both apoptosis and differentiating myoblasts and the conserved muscle contractile protein, myosin light chain kinase, is required for the apoptotic feature of membrane blebbing. As such, these observations suggest that the induction of differentiation and apoptosis in the myogenic lineage may use overlapping cellular mechanisms. Here, we report that skeletal muscle differentiation depends on the activity of the key apoptotic protease, caspase 3. Peptide inhibition of caspase 3 activity or homologous deletion of caspase 3 leads to dramatic reduction in both myotube/myofiber formation and expression of muscle-specific proteins. Subsequently, we have identified Mammalian Sterile Twenty-like kinase as a crucial caspase 3 effector in this cellular process. Mammalian Sterile Twenty-like kinase is cleavage-activated by caspase 3, and restoration of this truncated kinase in caspase 3 null myoblasts restores the differentiation phenotype. Taken together, these results confirm a unique and unanticipated role for a caspase 3-mediated signal cascade in the promotion of myogenesis.
Figures
References
-
- Megeney L. A. & Rudnicki, M. A. (1995) Biochem. Cell. Biol. 73, 723-732. - PubMed
-
- Black B. L. & Olson, E. N. (1998) Annu. Rev. Cell Dev. Biol. 14, 167-196. - PubMed
-
- Black B. L., Martin, J. F. & Olson, E. N. (1995) J. Biol. Chem. 270, 2889-2892. - PubMed
-
- Molkentin J. D., Black, B. L., Martin, J. F. & Olson, E. N. (1995) Cell 83, 1125-1136. - PubMed
-
- Yee S. P. & Rigby, P. W. (1993) Genes Dev. 7, 1277-1289. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
