

Regulation and role of the Arabidopsis abscisic acid-insensitive 5 gene in abscisic acid, sugar, and stress response
- PMID: 12177466
- PMCID: PMC166741
- DOI: 10.1104/pp.005793
Regulation and role of the Arabidopsis abscisic acid-insensitive 5 gene in abscisic acid, sugar, and stress response
Abstract
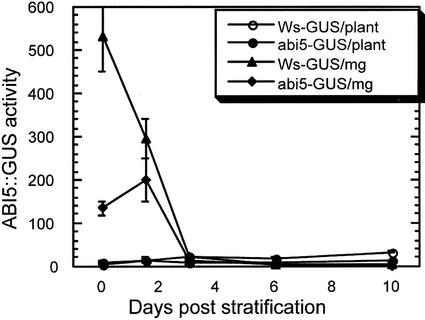
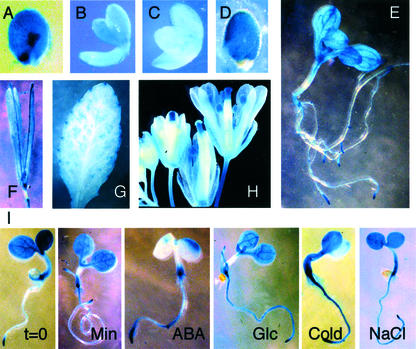
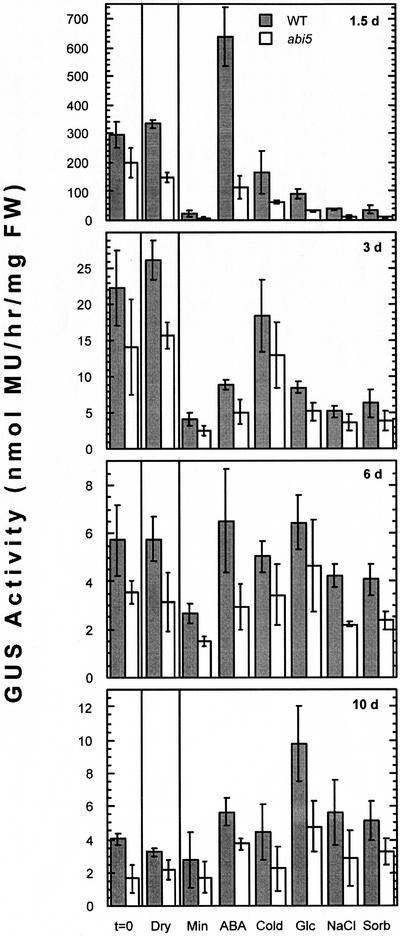
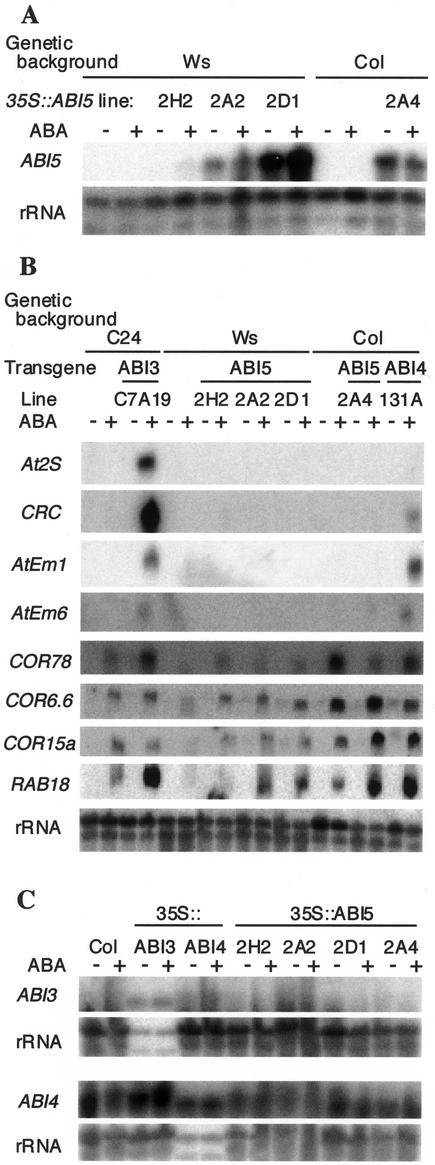
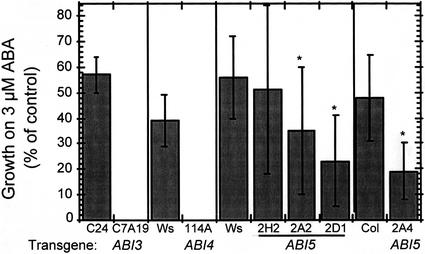
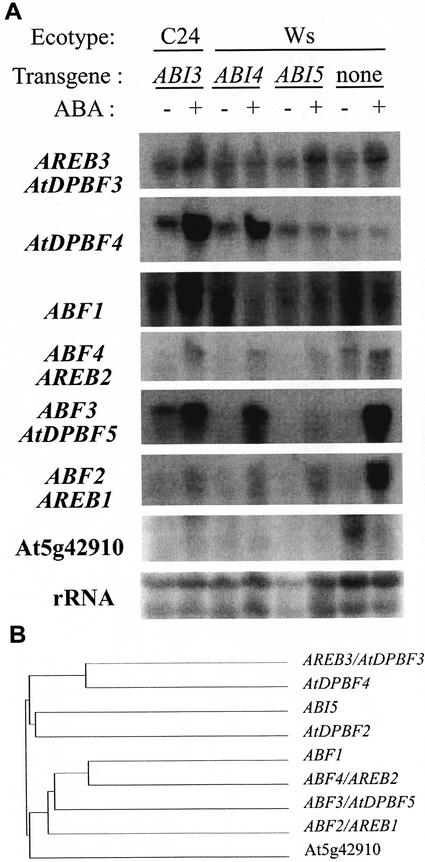
Abscisic acid (ABA) and stress response from late embryonic growth through early seedling development is regulated by a signaling network that includes the Arabidopsis ABA-insensitive (ABI)5 gene, which encodes a basic leucine zipper transcription factor. We have characterized genetic, developmental, and environmental regulation of ABI5 expression. Although expressed most strongly in seeds, the ABI5 promoter is also active in vegetative and floral tissue. Vegetative expression is strongly induced by ABA, and weakly by stress treatments during a limited developmental window up to approximately 2 d post-stratification, but ABA and some stresses can induce expression in specific tissues at later stages. ABI5 expression is autoregulated in transgenic plants and yeast (Saccharomyces cerevisiae), and stress response appears to involve ABI5-dependent and -independent mechanisms. To determine whether ABI5 is necessary and/or sufficient for ABA or stress response, we assayed the effects of increased ABI5 expression on growth and gene expression. Although overexpression of ABI5 confers hypersensitivity to ABA and sugar, as previously described for ABI4 and ABI3 overexpression lines, it has relatively limited effects on enhancing ABA-responsive gene expression. Comparison of expression of eight ABI5-homologous genes shows overlapping regulation by ABI3, ABI4, and ABI5, suggestive of a combinatorial network involving positive and negative regulatory interactions.
Figures


References
-
- An G, Ebert P, Mitra A, Ha S. Binary vectors. In: Gelvin S, Schilperoort R, editors. Plant Molecular Biology Manual. Dordrecht, The Netherlands: Kluwer Academic Publishers; 1988. , A3 pp 1–19.
-
- Bechtold N, Ellis J, Pelletier G. In planta Agrobacterium gene transfer by infiltration of adult Arabidopsis thaliana plants. C R Acad Sci Paris Life Sci. 1993;316:1194–1199.
-
- Chia DW, Yoder TJ, Reiter W-D, Gibson SI. Fumaric acid: an overlooked form of fixed carbon in Arabidopsis and other plant species. Planta. 2000;211:743–751. - PubMed
-
- Choi H, Hong J, Ha J, Kang J, Kim S. ABFs, a family of ABA-responsive element binding factors. J Biol Chem. 2000;275:1723–1730. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
