

Cadmium-induced sulfate uptake in maize roots
- PMID: 12177501
- PMCID: PMC166776
- DOI: 10.1104/pp.002659
Cadmium-induced sulfate uptake in maize roots
Abstract
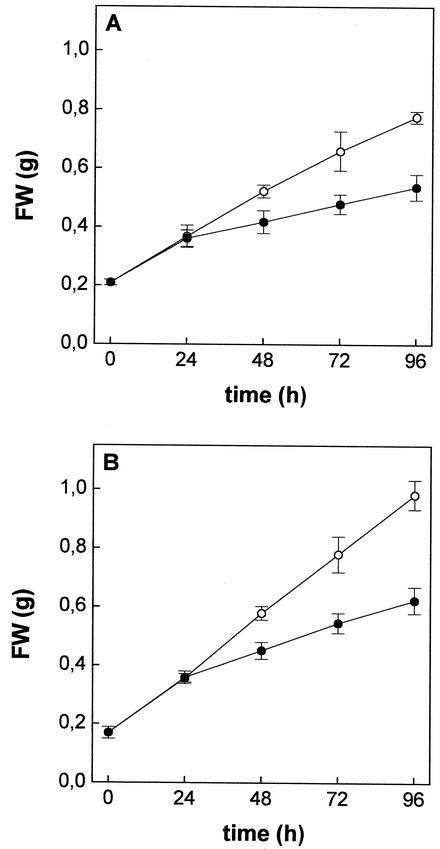
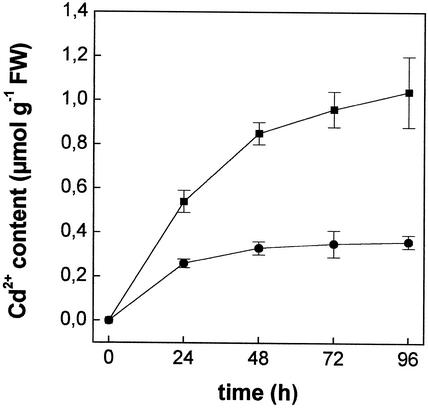
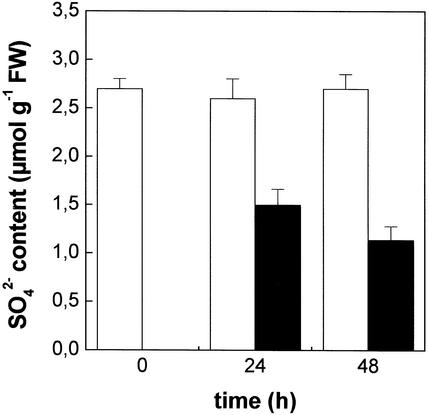
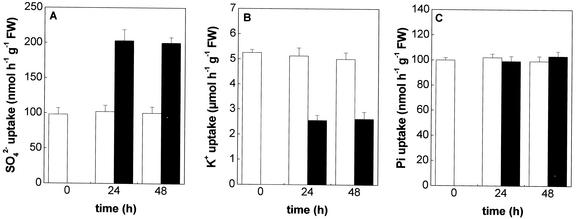
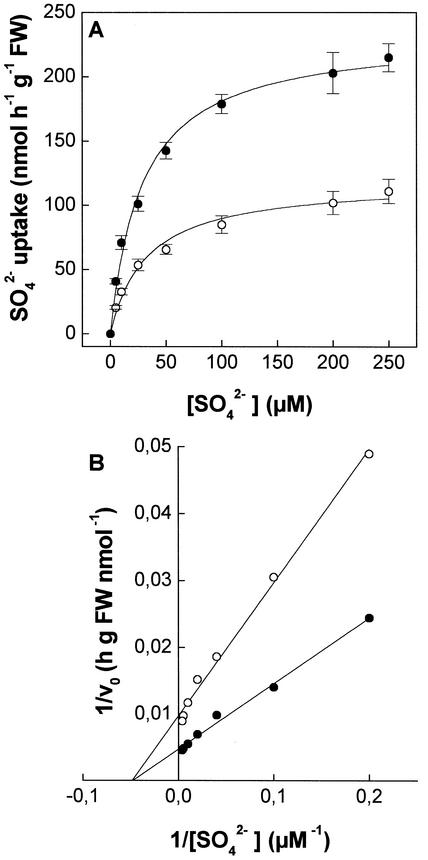
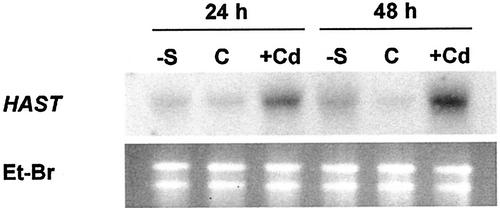
The effect of cadmium (Cd) on high-affinity sulfate transport of maize (Zea mays) roots was studied and related to the changes in the levels of sulfate and nonprotein thiols during Cd-induced phytochelatin (PC) biosynthesis. Ten micromolar CdCl(2) in the nutrient solution induced a 100% increase in sulfate uptake by roots. This was not observed either for potassium or phosphate uptake, suggesting a specific effect of Cd(2+) on sulfate transport. The higher sulfate uptake was not dependent on a change in the proton motive force that energizes it. In fact, in Cd-treated plants, the transmembrane electric potential difference of root cortical cells was only slightly more negative than in the controls, the external pH did not change, and the activity of the plasma membrane H(+)-ATPase did not increase. Kinetics analysis showed that in the range of the high-affinity sulfate transport systems, 10 to 250 microM, Cd exposure did not influence the K(m) value (about 20 microM), whereas it doubled the V(max) value with respect to the control. Northern-blot analysis showed that Cd-induced sulfate uptake was related to a higher level of mRNA encoding for a putative high-affinity sulfate transporter in roots. Cd-induced sulfate uptake was associated to both a decrease in the contents of sulfate and glutathione and synthesis of a large amount of PCs. These results suggest that Cd-induced sulfate uptake depends on a pretranslational regulation of the high-affinity sulfate transporter gene and that this response is necessary for sustaining the higher sulfur demand during PC biosynthesis.
Figures
References
-
- Bolchi A, Petrucco S, Tenca PL, Foroni C, Ottonello S. Coordinate modulation of maize sulfate permease and ATP sulfurylase mRNAs in response to variations in sulfur nutritional status: stereospecific down-regulation by l-cysteine. Plant Mol Biol. 1999;39:527–537. - PubMed
-
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. - PubMed
-
- Clarkson DT, Smith FW, Vanden Berg PJ. Regulation of sulphate transport in tropical legume, Macroptilum atropurpureum, cv. Siratro J Exp Bot. 1983;34:1463–1483.
-
- Clemens S. Molecular mechanisms of plant metal tolerance and homeostasis. Planta. 2001;212:475–486. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
