

The LisRK signal transduction system determines the sensitivity of Listeria monocytogenes to nisin and cephalosporins
- PMID: 12183229
- PMCID: PMC127401
- DOI: 10.1128/AAC.46.9.2784-2790.2002
The LisRK signal transduction system determines the sensitivity of Listeria monocytogenes to nisin and cephalosporins
Abstract
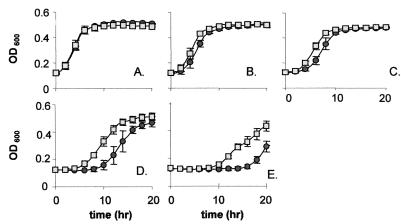
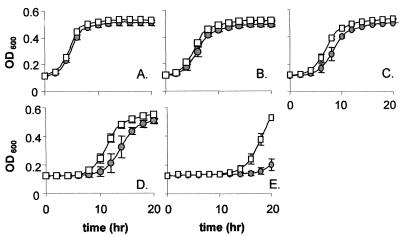
The Listeria monocytogenes two-component signal transduction system, LisRK, initially identified in strain LO28, plays a significant role in the virulence potential of this important food-borne pathogen. Here, it is shown that, in addition to its major contribution in responding to ethanol, pH, and hydrogen peroxide stresses, LisRK is involved in the ability of the cell to tolerate important antimicrobials used in food and in medicine, e.g., the lantibiotic nisin and the cephalosporin family of antibiotics. A (Delta)lisK mutant (lacking the LisK histidine kinase sensor component) displays significantly enhanced resistance to the lantibiotic nisin, a greatly enhanced sensitivity to the cephalosporins, and a large reduction in the expression of three genes thought to encode a penicillin-binding protein, another histidine kinase (other than LisK), and a protein of unknown function. Confirmation of the role of LisRK was obtained when the response regulator, LisR, was overexpressed using both constitutive and inducible (nisin-controlled expression) systems. Under these conditions we observed a reversion of the (Delta)lisK mutant to wild-type growth kinetics in the presence of nisin. It was also found that overexpression of LisR complemented the reduced expression of two of the aforementioned genes. These results demonstrate the important role of LisRK in the response of L. monocytogenes to a number of antimicrobial agents.
Figures

Similar articles
-
Assessing the contributions of the LiaS histidine kinase to the innate resistance of Listeria monocytogenes to nisin, cephalosporins, and disinfectants.Appl Environ Microbiol. 2012 Apr;78(8):2923-9. doi: 10.1128/AEM.07402-11. Epub 2012 Feb 10. Appl Environ Microbiol. 2012. PMID: 22327581 Free PMC article.
-
Gene expression profiling of a nisin-sensitive Listeria monocytogenes Scott A ctsR deletion mutant.J Ind Microbiol Biotechnol. 2013 May;40(5):495-505. doi: 10.1007/s10295-013-1243-0. Epub 2013 Mar 14. J Ind Microbiol Biotechnol. 2013. PMID: 23494707
-
Spontaneous nisin-resistant Listeria monocytogenes mutants with increased expression of a putative penicillin-binding protein and their sensitivity to various antibiotics.Microb Drug Resist. 2001 Summer;7(2):127-35. doi: 10.1089/10766290152045002. Microb Drug Resist. 2001. PMID: 11442339
-
Nisin resistance of Listeria monocytogenes is increased by exposure to salt stress and is mediated via LiaR.Appl Environ Microbiol. 2013 Sep;79(18):5682-8. doi: 10.1128/AEM.01797-13. Epub 2013 Jul 12. Appl Environ Microbiol. 2013. PMID: 23851083 Free PMC article.
-
The intrinsic cephalosporin resistome of Listeria monocytogenes in the context of stress response, gene regulation, pathogenesis and therapeutics.J Appl Microbiol. 2016 Feb;120(2):251-65. doi: 10.1111/jam.12989. Epub 2015 Dec 28. J Appl Microbiol. 2016. PMID: 26509460 Review.
Cited by
-
Comprehensive studies of drug resistance mediated by overexpression of response regulators of two-component signal transduction systems in Escherichia coli.J Bacteriol. 2003 Mar;185(6):1851-6. doi: 10.1128/JB.185.6.1851-1856.2003. J Bacteriol. 2003. PMID: 12618449 Free PMC article.
-
Cold Shock Proteins Promote Nisin Tolerance in Listeria monocytogenes Through Modulation of Cell Envelope Modification Responses.Front Microbiol. 2021 Dec 24;12:811939. doi: 10.3389/fmicb.2021.811939. eCollection 2021. Front Microbiol. 2021. PMID: 35003042 Free PMC article.
-
Tools for functional postgenomic analysis of listeria monocytogenes.Appl Environ Microbiol. 2008 Jul;74(13):3921-34. doi: 10.1128/AEM.00314-08. Epub 2008 Apr 25. Appl Environ Microbiol. 2008. PMID: 18441118 Free PMC article.
-
Comparison of the Potency of the Lipid II Targeting Antimicrobials Nisin, Lacticin 3147 and Vancomycin Against Gram-Positive Bacteria.Probiotics Antimicrob Proteins. 2012 Jun;4(2):108-15. doi: 10.1007/s12602-012-9095-x. Probiotics Antimicrob Proteins. 2012. PMID: 26781852
-
The multicopy sRNA LhrC controls expression of the oligopeptide-binding protein OppA in Listeria monocytogenes.RNA Biol. 2015;12(9):985-97. doi: 10.1080/15476286.2015.1071011. RNA Biol. 2015. PMID: 26176322 Free PMC article.
References
-
- Breukink, E., C. van Kraaij, R. A. Demel, R. J. Siezen, O. P. Kuipers, and B. de Kruijff. 1997. The C-terminal region of nisin is responsible for the initial interaction of nisin with the target membrane. Biochemistry 36:6968-6976. - PubMed
-
- Breukink, E., I. Wiedmann, C. van Kraaij, O. P. Kuipers, H. Sahl, and B. de Kruijff. 1999. Use of the cell wall precursor lipid II by a pore-forming peptide antibiotic. Science 286:2361-2364. - PubMed
-
- Bryan, E. M., T. Bae, M. Kleerebezem, and G. M. Dunny. 2000. Improved vectors for nisin-controlled expression in gram-positive bacteria. Plasmid 44:183-190. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
