

Samarangenin B from Limonium sinense suppresses herpes simplex virus type 1 replication in Vero cells by regulation of viral macromolecular synthesis
- PMID: 12183238
- PMCID: PMC127446
- DOI: 10.1128/AAC.46.9.2854-2864.2002
Samarangenin B from Limonium sinense suppresses herpes simplex virus type 1 replication in Vero cells by regulation of viral macromolecular synthesis
Abstract
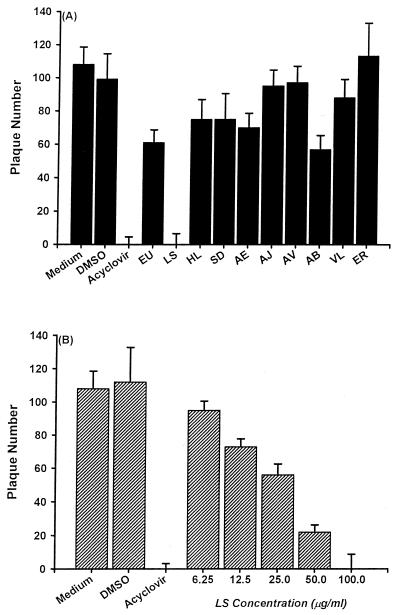
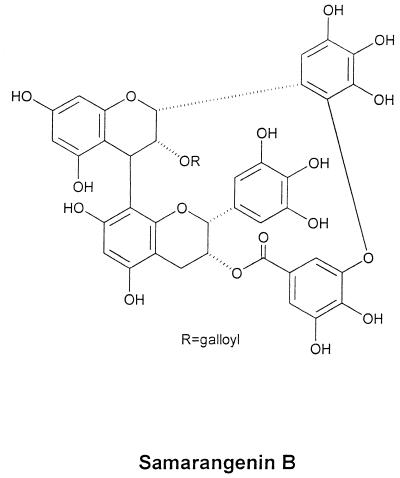
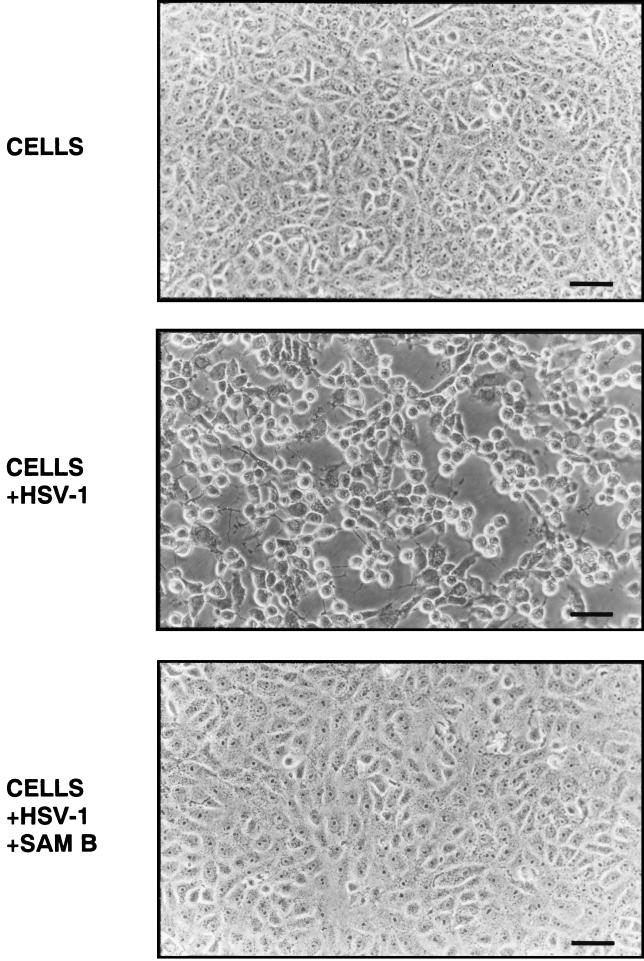
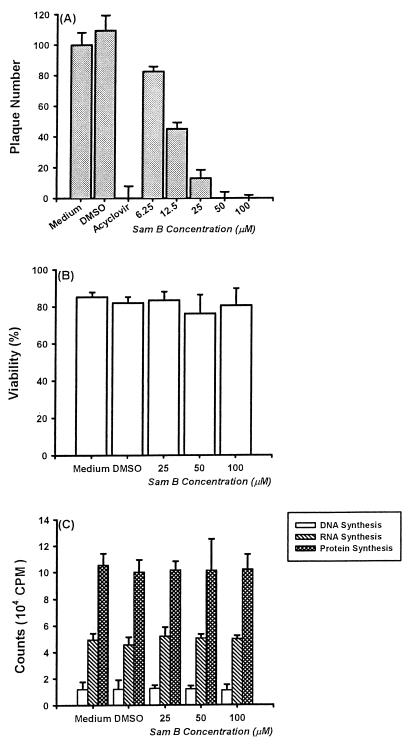
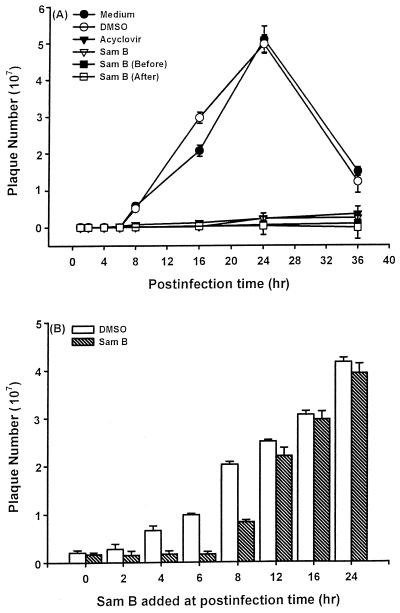
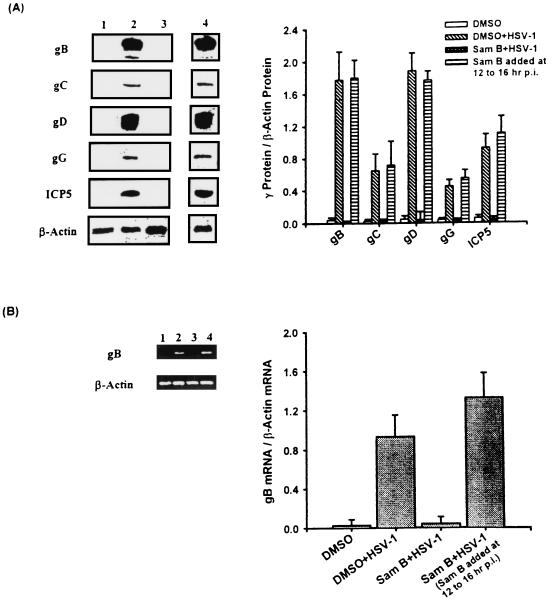
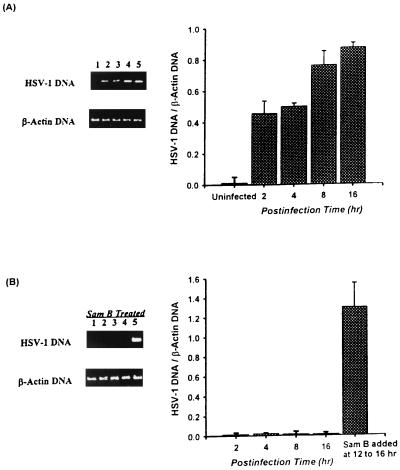
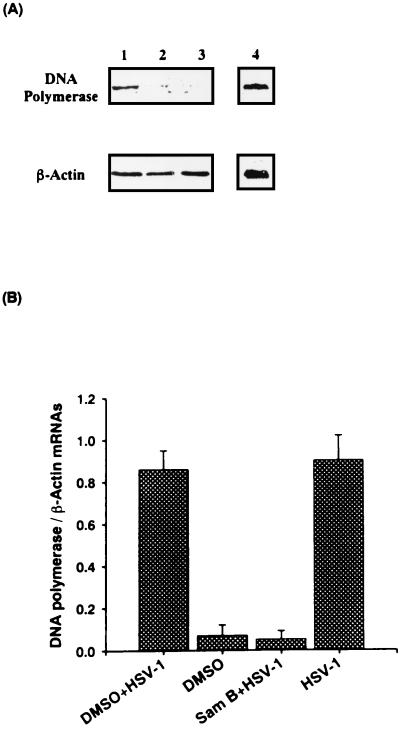
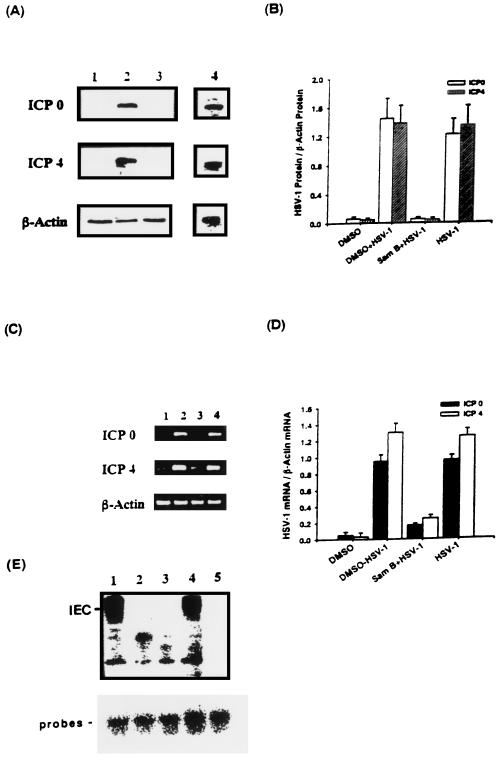
Inhibitory effects of ethanolic extracts from 10 Chinese herbs on herpes simplex virus type 1 (HSV-1) replication were investigated. By a bioassay-guided fractionation procedure, samarangenin B (Sam B) was isolated from Limonium sinense; Sam B significantly suppressed HSV-1 multiplication in Vero cells without apparent cytotoxicity. Time-of-addition experiments suggested that the inhibitory action of Sam B on HSV-1 replication was not due to the blocking of virus adsorption. In an attempt to further localize the point in the HSV-1 replication cycle where arrest occurred, a set of key regulatory events leading to viral multiplication was examined, including viral immediate-early (alpha), early (beta), and late (gamma) gene expression and DNA replication. Results indicated that levels of glycoprotein B (gB), gC, gD, gG, and infected-cell protein 5 (ICP5) expression and gB mRNA expression in Vero cells were impeded by Sam B. Data from PCR showed that replication of HSV-1 DNA in Vero cells was arrested by Sam B. Furthermore, Sam B decreased DNA polymerase, ICP0, and ICP4 gene expression in Vero cells. Results of an electrophoretic mobility shift assay demonstrated that Sam B interrupted the formation of an alpha-trans-induction factor/C1/Oct-1/GARAT multiprotein complex. The mechanisms of antiviral action of Sam B seem to be mediated, at least in part, by inhibiting HSV-1 alpha gene expression, including expression of the ICP0 and ICP4 genes, by blocking beta transcripts such as DNA polymerase mRNA, and by arresting HSV-1 DNA synthesis and structural protein expression in Vero cells. These results show that Sam B is an antiviral agent against HSV-1 replication.
Figures
Similar articles
-
Yatein from Chamaecyparis obtusa suppresses herpes simplex virus type 1 replication in HeLa cells by interruption the immediate-early gene expression.Antiviral Res. 2006 Jul;70(3):112-20. doi: 10.1016/j.antiviral.2006.01.011. Epub 2006 Feb 20. Antiviral Res. 2006. PMID: 16540181
-
Regulation of herpes simplex virus type 1 replication in Vero cells by Psychotria serpens: relationship to gene expression, DNA replication, and protein synthesis.Antiviral Res. 2001 Aug;51(2):95-109. doi: 10.1016/s0166-3542(01)00141-3. Antiviral Res. 2001. PMID: 11431035
-
Herpes simplex virus type 1 propagation in HeLa cells interrupted by Nelumbo nucifera.J Biomed Sci. 2005 Dec;12(6):1021-34. doi: 10.1007/s11373-005-9001-6. Epub 2005 Aug 18. J Biomed Sci. 2005. PMID: 16132118
-
Resveratrol suppresses nuclear factor-kappaB in herpes simplex virus infected cells.Antiviral Res. 2006 Dec;72(3):242-51. doi: 10.1016/j.antiviral.2006.06.011. Epub 2006 Jul 14. Antiviral Res. 2006. PMID: 16876885
-
The immediate-early protein, ICP0, is essential for the resistance of herpes simplex virus to interferon-alpha/beta.Virology. 2002 Feb 15;293(2):295-304. doi: 10.1006/viro.2001.1280. Virology. 2002. PMID: 11886249
Cited by
-
Sulfated phenolic compounds from Limonium caspium: Isolation, structural elucidation, and biological evaluation.Fitoterapia. 2015 Jul;104:80-5. doi: 10.1016/j.fitote.2015.05.017. Epub 2015 May 27. Fitoterapia. 2015. PMID: 26025854 Free PMC article.
-
Traditional Chinese herbal medicine as a source of molecules with antiviral activity.Antiviral Res. 2013 Jan;97(1):1-9. doi: 10.1016/j.antiviral.2012.10.006. Epub 2012 Nov 12. Antiviral Res. 2013. PMID: 23153834 Free PMC article. Review.
-
Bio-guided chemical characterization and nano-formulation studies of selected edible volatile oils with potentials antibacterial and anti-SARS-CoV-2 activities.Arab J Chem. 2023 Jul;16(7):104813. doi: 10.1016/j.arabjc.2023.104813. Epub 2023 Mar 18. Arab J Chem. 2023. PMID: 36969951 Free PMC article.
-
Chemical constituents from Limonium tubiflorum and their in silico evaluation as potential antiviral agents against SARS-CoV-2.RSC Adv. 2021 Sep 30;11(51):32346-32357. doi: 10.1039/d1ra05927k. eCollection 2021 Sep 27. RSC Adv. 2021. PMID: 35495487 Free PMC article.
-
Integrated network pharmacology and cellular assay reveal the biological mechanisms of Limonium sinense (Girard) Kuntze against Breast cancer.BMC Complement Med Ther. 2023 Nov 13;23(1):408. doi: 10.1186/s12906-023-04233-z. BMC Complement Med Ther. 2023. PMID: 37957642 Free PMC article.
References
-
- Alrabiah, F. A., and S. L. Sacks. 1996. New antiherpes virus agents: their targets and therapeutic potential. Drugs 52:17-32. - PubMed
-
- Arisawa, M., A. Fujita, T. Hayashi, K. Hayashi, H. Ochiai, and N. Morita. 1990. Cytotoxic and antiherpetic activity of phloroglucinol derivatives from Mallotus japonicus (Euphorbiaceae). Chem. Pharm. Bull. 38:1624-1626. - PubMed
-
- Bacon, T. H., B. A. Howard, L. C. Spender, and M. R. Boyd. 1996. Activity of penciclovir in antiviral assays against herpes simplex virus. J. Antimicrob. Chemother. 37:303-313. - PubMed
-
- Boulware, S. L., J. C. Bronstein, E. C. Nordby, and P. C. Weber. 2001. Identification and characterization of a benzothiophene inhibitor of herpes simplex virus type 1 replication which acts at the immediate early stage of infection. Antivir. Res. 51:111-125. - PubMed
-
- Chen, H., L. Teng, J. N. Li, R. Park, D. E. Mold, J. Gnabre, J. R. Hwu, W. N. Tseng, and R. C. Huang. 1998. Antiviral activities of methylated nordihydroguaiaretic acids. 2. Targeting herpes simplex virus replication by the mutation insensitive transcription inhibitor tetra-O-methyl-NDGA. J. Med. Chem. 41:3001-3007. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
