

Induction of the gene encoding macrophage chemoattractant protein 1 by Orientia tsutsugamushi in human endothelial cells involves activation of transcription factor activator protein 1
- PMID: 12183528
- PMCID: PMC128290
- DOI: 10.1128/IAI.70.9.4841-4850.2002
Induction of the gene encoding macrophage chemoattractant protein 1 by Orientia tsutsugamushi in human endothelial cells involves activation of transcription factor activator protein 1
Abstract
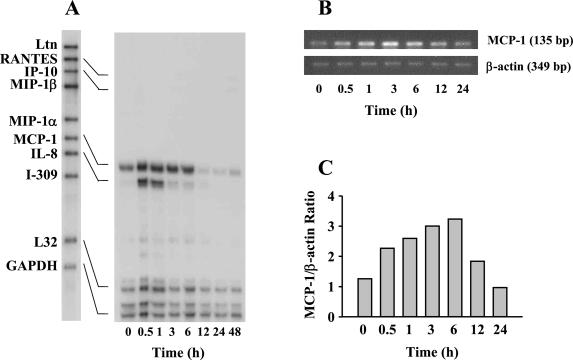
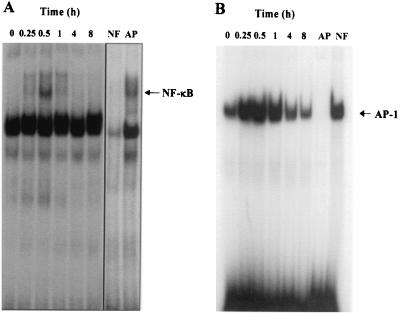
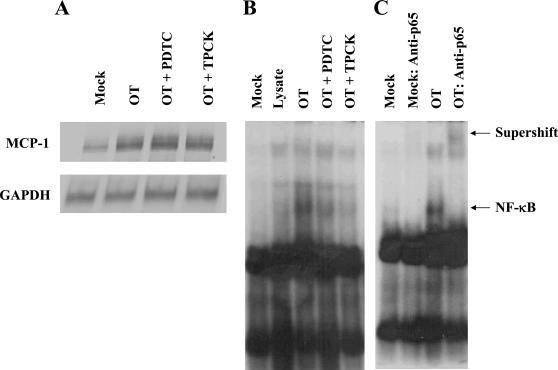
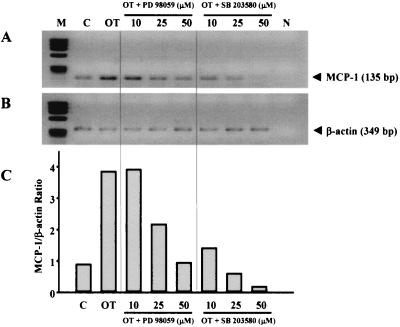
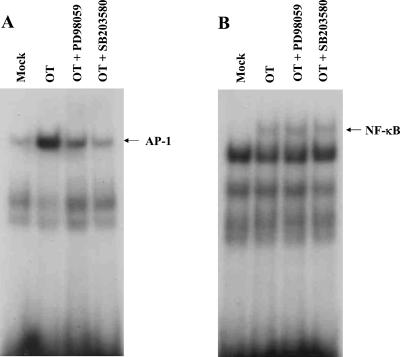
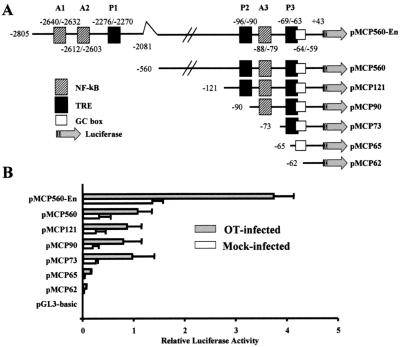
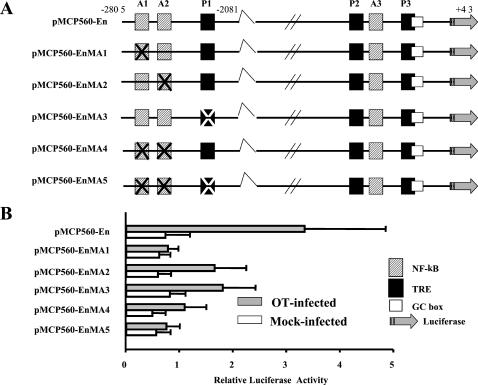
Human macrophage chemoattractant protein 1 (MCP-1) is a potent mediator of macrophage migration and therefore plays an essential role in early events of inflammation. In endothelial cells, at least three independent pathways regulate MCP-1 expression by NF-kappaB and AP-1. Orientia tsutsugamushi causes vasculitis in humans by replicating inside macrophages and endothelial cells. In the present study, we investigated the cis-acting and trans-acting elements involved in O. tsutsugamushi-induced MCP-1 gene expression in human umbilical vein endothelial cells (HUVEC). Although NF-kappaB activation was observed in HUVEC infected with O. tsutsugamushi, inhibition of NF-kappaB activation did not affect the MCP-1 expression. However, treatment of HUVEC with extracellular signal-regulated kinase (ERK) kinase inhibitor or p38 mitogen-activated protein kinase (MAPK) inhibitor suppressed expression of MCP-1 mRNA concomitant with downregulation of activator protein 1 (AP-1) activation. Deletion of triphorbol acetate response elements (TRE) at position -69 to -63 of MCP-1 gene abolished inducible promoter activity. Deletion of TRE at position -69 to -63-96 to -90 or deletion of NF-kappaB-binding site at position -69 to -63-88 to -79 did not affect the inducibility of promoter. Site-directed mutagenesis of the NF-kappaB binding sites at positions -2640 to -2632, -2612 to -2603 in the enhancer region, or the AP-1 biding site at position -2276 to -2270 decreased the inducible activity of the promoter. Taken together, AP-1 activation by both the ERK pathway and the p38 MAPK pathway as well as their binding to TRE at position -69 to -63 in proximal promoter and TRE at position -2276 to -2270 in enhancer region is altogether essential in induction of MCP-1 mRNA in HUVEC infected with O. tsutsugamushi. Although NF-kappaB activation is not essential per se, the kappaB site in the enhancer region is important in MCP-1 induction of HUVEC. This discrepancy in the involvement of the NF-kappaB may be due to the function of chromatin structures and other transcription cofactors in the regulation of MCP-1 gene expression in response to O. tsutsugamushi infectioin.
Figures
References
-
- Baggiolini, M., B. Dewald, and B. Moser. 1994. Interleukin-8 and related chemotactic cytokines-CXC and CC chemokines. Adv. Immunol. 55:97-179. - PubMed
-
- Baldwin, A. S., Jr. 1996. The NF-kappa B and I kappa B proteins: new discoveries and insights. Annu. Rev. Immunol. 14:649-683. - PubMed
-
- Beilke, M. A. 1989. Vascular endothelium in immunology and infectious disease. Rev. Infect. Dis. 11:273-283. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
