

Incorporation of N-acetylneuraminic acid into Haemophilus somnus lipooligosaccharide (LOS): enhancement of resistance to serum and reduction of LOS antibody binding
- PMID: 12183531
- PMCID: PMC128230
- DOI: 10.1128/IAI.70.9.4870-4879.2002
Incorporation of N-acetylneuraminic acid into Haemophilus somnus lipooligosaccharide (LOS): enhancement of resistance to serum and reduction of LOS antibody binding
Erratum in
- Infect Immun 2002 Nov;70(11):6512
Abstract
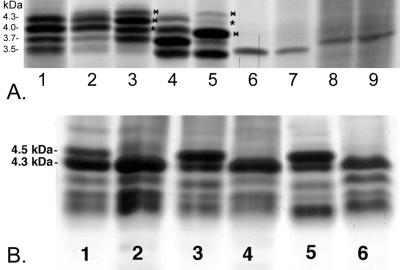
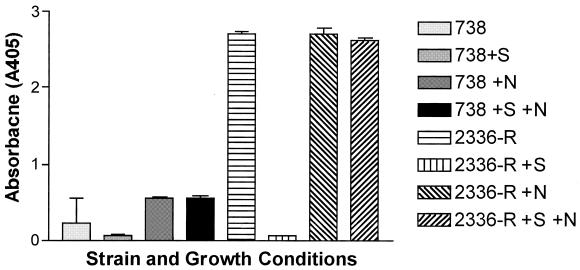
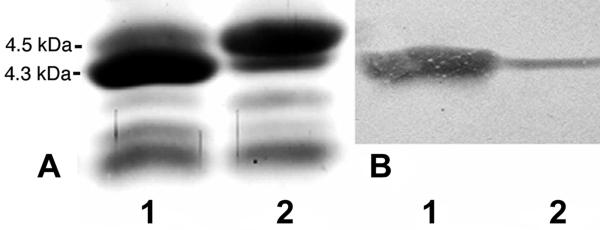
Haemophilus somnus isolates from cases of thrombotic meningoencephalitis, pneumonia, and other disease sites are capable of undergoing a high rate of phase variation in the oligosaccharide component of their lipooligosaccharides (LOS). In contrast, the LOS of commensal strains isolated from the normal reproductive tract phase vary little or not at all. In addition, the LOS of H. somnus shares conserved epitopes with LOS from Neisseria gonorrhoeae, Haemophilus influenzae, and other species that can incorporate sialic acid into their LOS. We now report that growth of disease isolates of H. somnus with CMP-N-acetylneuraminic acid (CMP-NeuAc) or NeuAc added to the medium resulted in incorporation of NeuAc into the LOS. However, NeuAc was not incorporated into the LOS of commensal isolates and one disease isolate following growth in medium containing CMP-NeuAc or NeuAc. Sialylated LOS was detected by an increase in the molecular size or an increase in the amount of the largest-molecular-size LOS electrophoretic bands, which disappeared following treatment with neuraminidase. Sialylated LOS could also be detected by reactivity with Limax flavus agglutinin lectin, which is specific for sialylated species, by dot blot assay; this reactivity was also reversed by neuraminidase treatment. H. somnus strain 2336 LOS was found to contain some sialic acid when grown in medium lacking CMP-NeuAc or NeuAc, although supplementation enhanced NeuAc incorporation. In contrast strain 738, an LOS phase variant of strain 2336, was less extensively sialylated when the growth medium was supplemented with CMP-NeuAc or NeuAc, as determined by electrophoretic profiles and electrospray mass spectrometry. The sialyltransferase of H. somnus strain 738 was confirmed to preferentially sialylate the Gal(beta)-(1-3)-GlcNAc component of the lacto-N-tetraose structure by capillary electrophoresis assay. Enhanced sialylation of the strain 2336 LOS inhibited the binding of monoclonal antibodies to LOS by enzyme immunoassay and Western blotting. Furthermore, sialylation of the LOS enhanced the resistance of H. somnus to the bactericidal action of antiserum to LOS. Sialylation and increased resistance to killing by normal serum also occurred in a deletion mutant that was deficient in the terminal Gal-GlcNAc disaccharide. LOS sialylation may therefore be an important virulence mechanism to protect H. somnus against the host immune system.
Figures


References
-
- Cole, S. P., D. G. Guiney, and L. B. Corbeil. 1992. Two linked genes for outer membrane proteins are absent in four non-disease strains of Haemophilus somnus. Mol. Microbiol. 6:1895-1902. - PubMed
-
- Corbeil, L. B., R. P. Gogolewski, L. R. Stephens, and T. J. Inzana. 1995. Haemophilus somnus: antigen analysis and immune responses, p. 63-73. In W. Donachie, F. A. Lainson, and J. C. Hodgson (ed.), Haemophilus, Actinobacillus, and Pasteurella. Plenum Press, New York, N.Y.
-
- Cox, A. D., M. D. Howard, J.-R. Brisson, M. Van Der Zwan, P. Thibault, M. B. Perry, and T. J. Inzana. 1998. Structural analysis of the phase-variable lipooligosaccharide from Haemophilus somnus strain 738. Eur. J. Biochem. 253:507-516. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
