

Inhibition of allogeneic T cell proliferation by indoleamine 2,3-dioxygenase-expressing dendritic cells: mediation of suppression by tryptophan metabolites
- PMID: 12186837
- PMCID: PMC2196057
- DOI: 10.1084/jem.20020052
Inhibition of allogeneic T cell proliferation by indoleamine 2,3-dioxygenase-expressing dendritic cells: mediation of suppression by tryptophan metabolites
Abstract
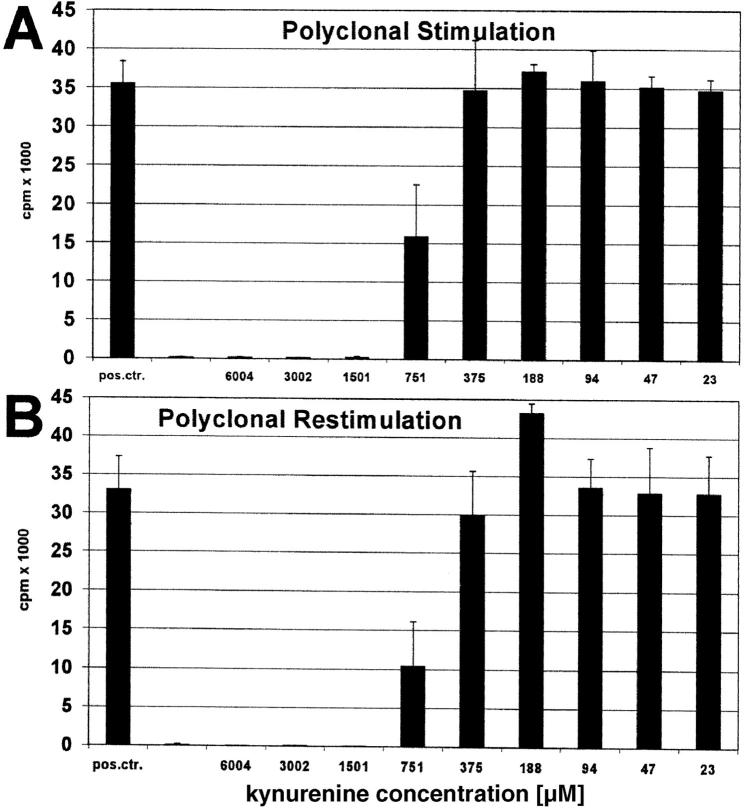
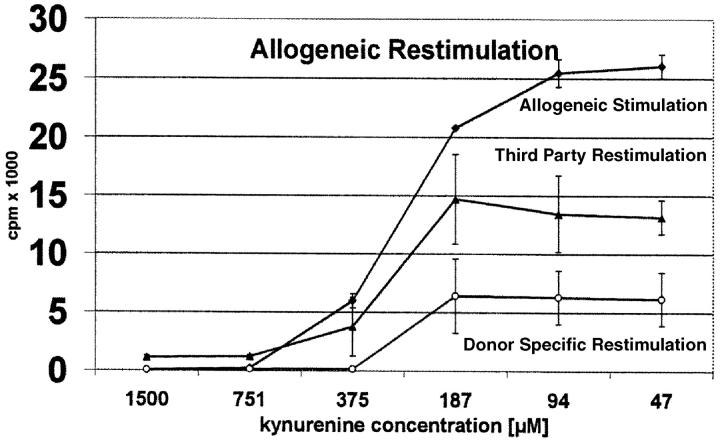
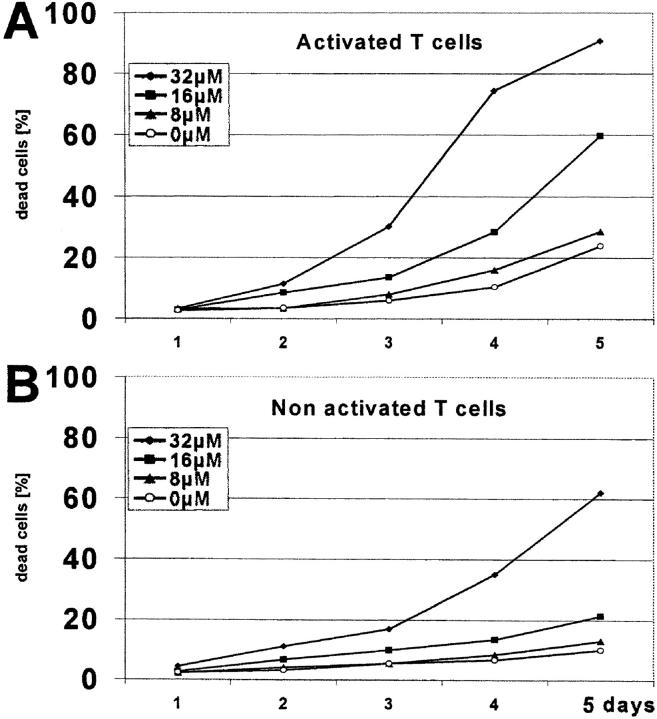
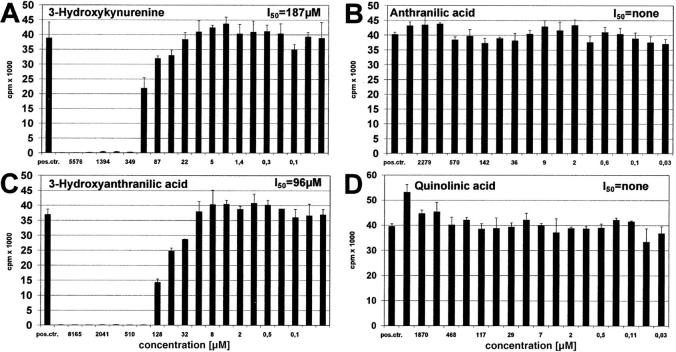
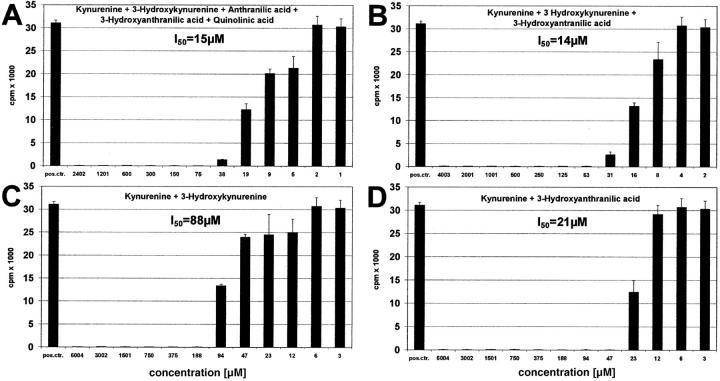
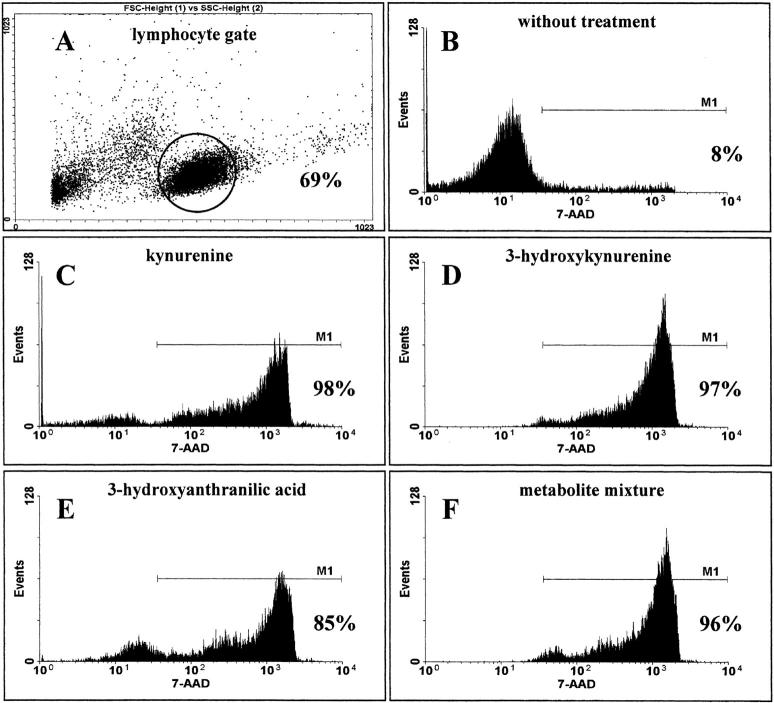
Indoleamine 2,3-dioxygenase (IDO), an enzyme involved in the catabolism of tryptophan, is expressed in certain cells and tissues, particularly in antigen-presenting cells of lymphoid organs and in the placenta. It was shown that IDO prevents rejection of the fetus during pregnancy, probably by inhibiting alloreactive T cells, and it was suggested that IDO-expression in antigen-presenting cells may control autoreactive immune responses. Degradation of tryptophan, an essential amino acid required for cell proliferation, was reported to be the mechanism of IDO-induced T cell suppression. Because we wanted to study the action of IDO-expressing dendritic cells (DCs) on allogeneic T cells, the human IDO gene was inserted into an adenoviral vector and expressed in DCs. Transgenic DCs decreased the concentration of tryptophan, increased the concentration of kynurenine, the main tryptophan metabolite, and suppressed allogeneic T cell proliferation in vitro. Kynurenine, 3-hydroxykynurenine, and 3-hydroxyanthranilic acid, but no other IDO-induced tryptophan metabolites, suppressed the T cell response, the suppressive effects being additive. T cells, once stopped in their proliferation, could not be restimulated. Inhibition of proliferation was likely due to T cell death because suppressive tryptophan catabolites exerted a cytotoxic action on CD3(+) cells. This action preferentially affected activated T cells and increased gradually with exposure time. In addition to T cells, B and natural killer (NK) cells were also killed, whereas DCs were not affected. Our findings shed light on suppressive mechanisms mediated by DCs and provide an explanation for important biological processes in which IDO activity apparently is increased, such as protection of the fetus from rejection during pregnancy and possibly T cell death in HIV-infected patients.
Figures
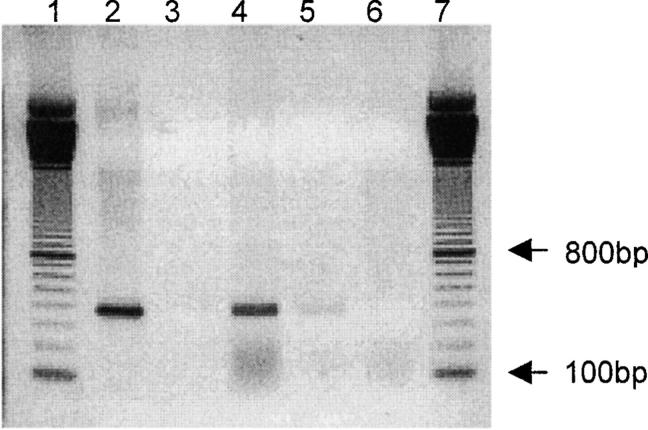
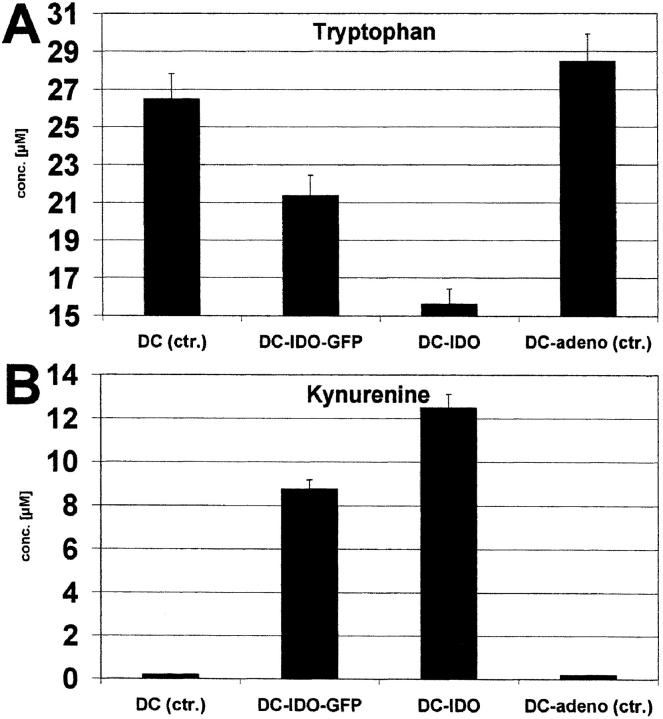
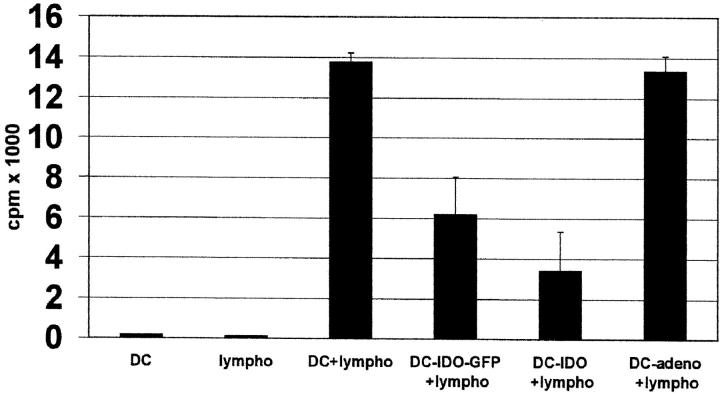
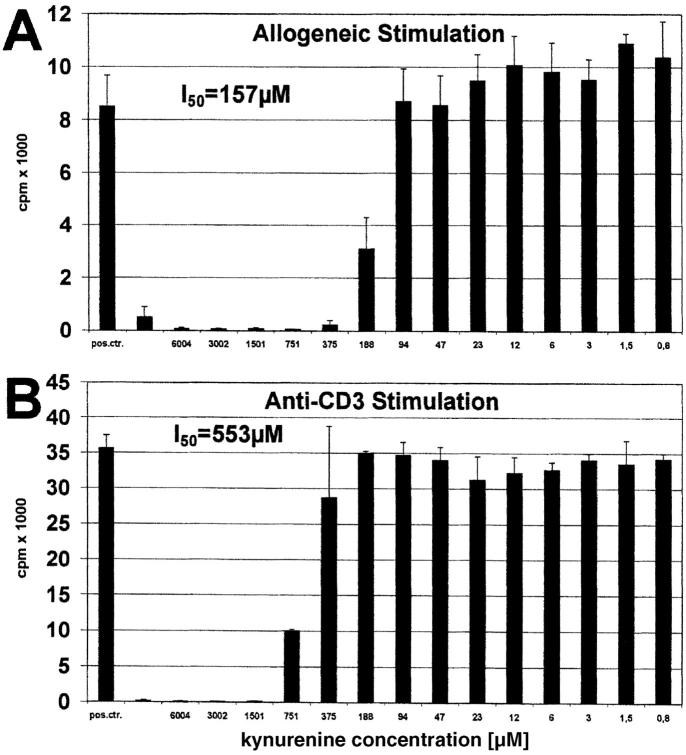

References
-
- Yoshida, R., T. Nukiwa, Y. Watanabe, M. Fujiwara, F. Hirata, and O. Hayashi. 1980. Regulation of indoleamine 2,3-dioxygenase activity in the small intestine and the epididymis of mice. Arch. Biochem. Biophys. 203:343–351. - PubMed
-
- Taylor, M.W., and G. Feng. 1991. Relationship between interferon-γ, indoleamine 2,3-dioxygenase, and tryptophan catabolism. FASEB J. 5:2516–2522. - PubMed
-
- Mellor, A.L., and D.H. Munn. 1999. Tryptophan catabolism and T-cell tolerance: immunosuppression by starvation? Immunol. Today. 20:469–473. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
