

A survival pathway for Caenorhabditis elegans with a blocked unfolded protein response
- PMID: 12186849
- PMCID: PMC2174003
- DOI: 10.1083/jcb.200203086
A survival pathway for Caenorhabditis elegans with a blocked unfolded protein response
Abstract
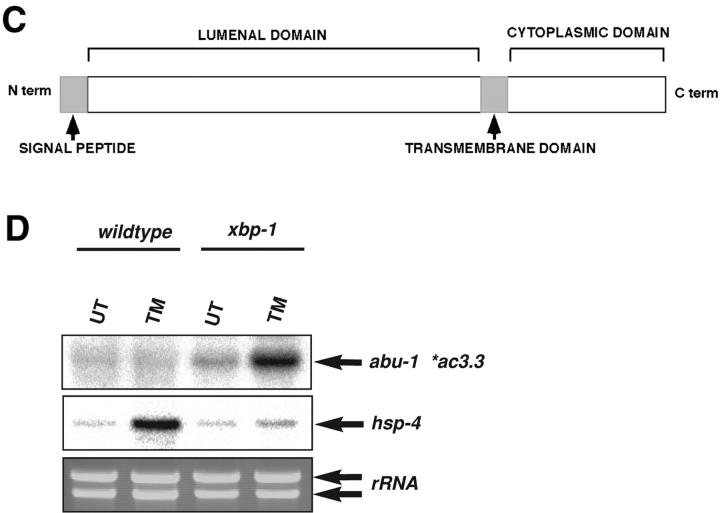
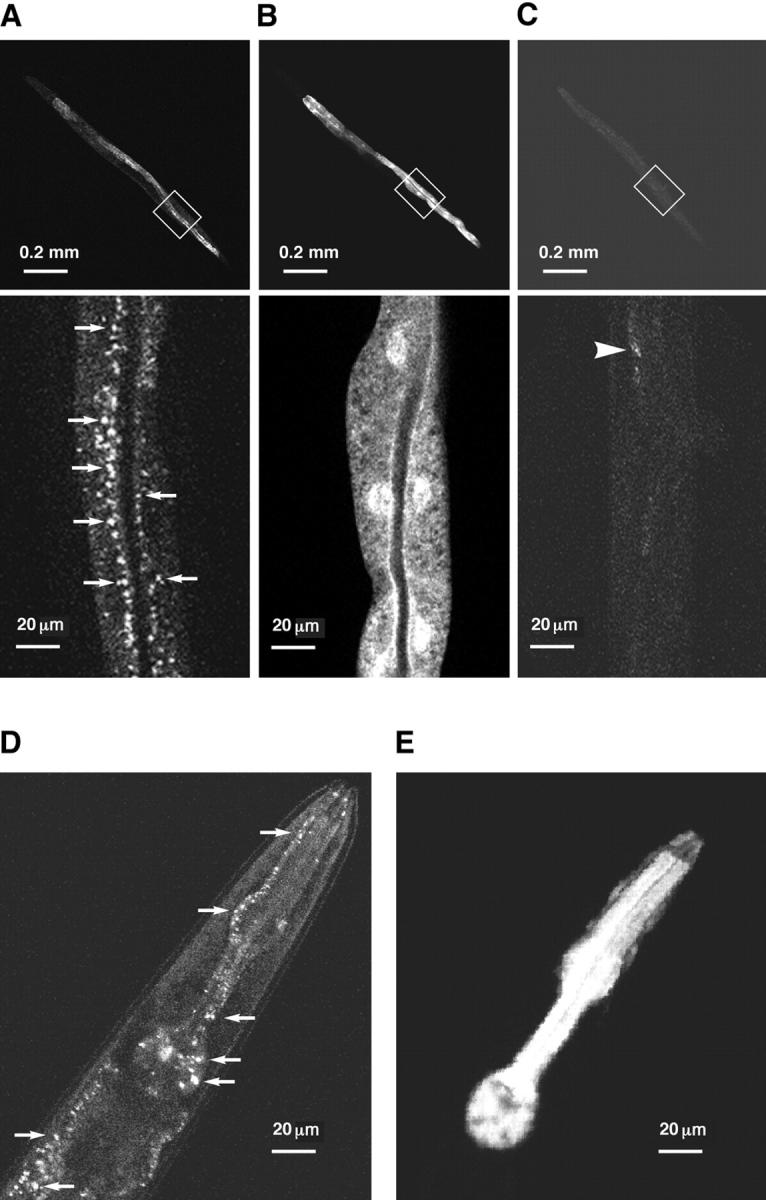
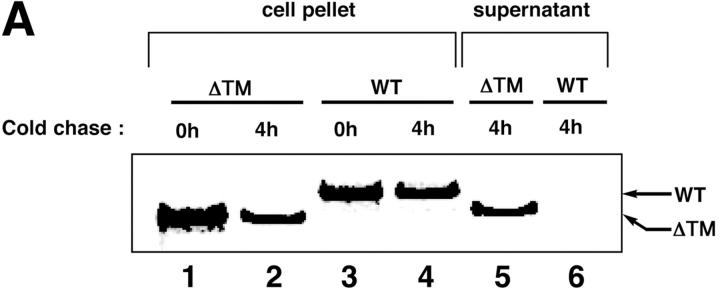
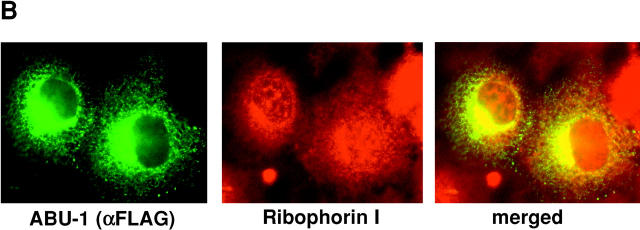
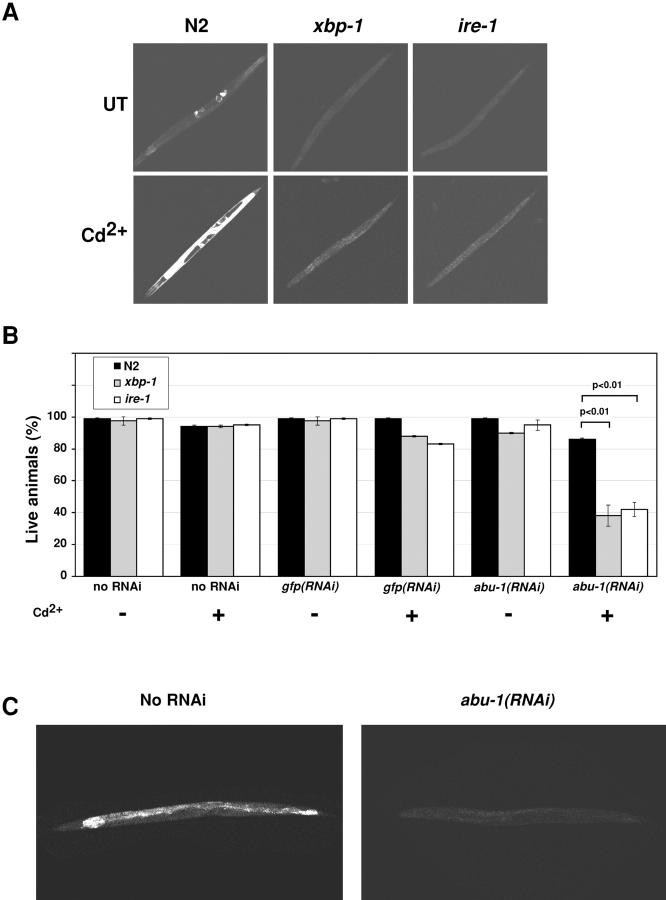
The unfolded protein response (UPR) counteracts stress caused by unprocessed ER client proteins. A genome-wide survey showed impaired induction of many UPR target genes in xbp-1 mutant Caenorhabditis elegans that are unable to signal in the highly conserved IRE1-dependent UPR pathway. However a family of genes, abu (activated in blocked UPR), was induced to higher levels in ER-stressed xbp-1 mutant animals than in ER-stressed wild-type animals. RNA-mediated interference (RNAi) inactivation of a representative abu family member, abu-1 (AC3.3), activated the ER stress marker hsp-4::gfp in otherwise normal animals and killed 50% of ER-stressed ire-1 and xbp-1 mutant animals. Abu-1(RNAi) also enhanced the effect of inactivation of sel-1, an ER-associated protein degradation gene. The nine abu genes encode highly related type I transmembrane proteins whose lumenal domains have sequence similarity to a mammalian cell surface scavenger receptor of endothelial cells that binds chemically modified extracellular proteins and directs their lysosomal degradation. Our findings that ABU-1 is an intracellular protein located within the endomembrane system that is induced by ER stress in xbp-1 mutant animals suggest that ABU proteins may interact with abnormal ER client proteins and this function may be particularly important in animals with an impaired UPR.
Figures


References
-
- Adachi, H., M. Tsujimoto, H. Arai, and K. Inoue. 1997. Expression cloning of a novel scavenger receptor from human endothelial cells. J. Biol. Chem. 272:31217–31220. - PubMed
-
- Aridor, M., and W.E. Balch. 1999. Integration of endoplasmic reticulum signaling in health and disease. Nat. Med. 5:745–751. - PubMed
-
- Calfon, M., H. Zeng, F. Urano, J.H. Till, S.R. Hubbard, H.P. Harding, S.G. Clark, and D. Ron. 2002. IRE1 couples endoplasmic reticulum load to secretory capacity by processing the XBP-1 mRNA. Nature. 415:92–96. - PubMed
-
- Carrell, R.W., and D.A. Lomas. 1997. Conformational disease. Lancet. 350:134–138. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
