

Effect of altering the tRNA(Lys)(3) concentration in human immunodeficiency virus type 1 upon its annealing to viral RNA, GagPol incorporation, and viral infectivity
- PMID: 12186893
- PMCID: PMC136429
- DOI: 10.1128/jvi.76.18.9096-9102.2002
Effect of altering the tRNA(Lys)(3) concentration in human immunodeficiency virus type 1 upon its annealing to viral RNA, GagPol incorporation, and viral infectivity
Abstract
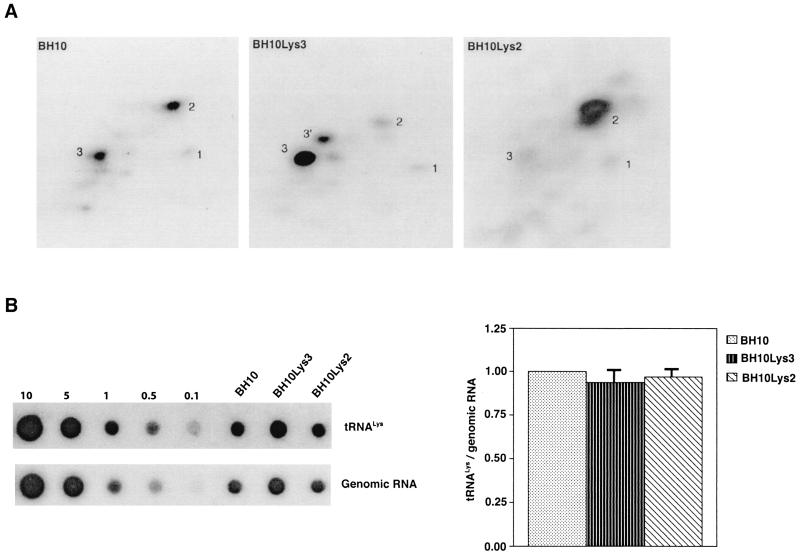
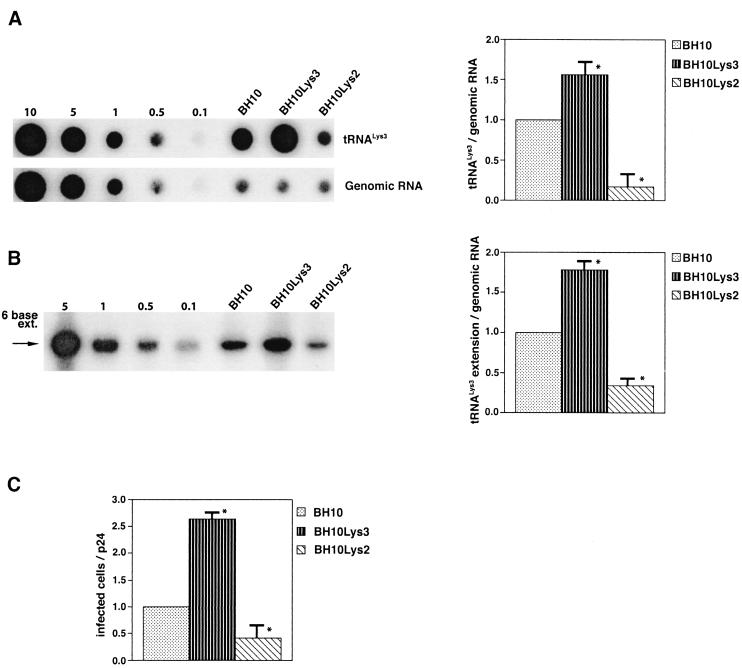
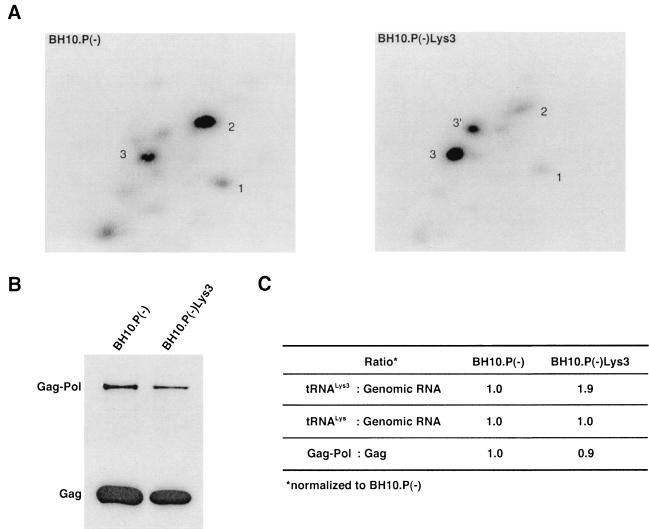
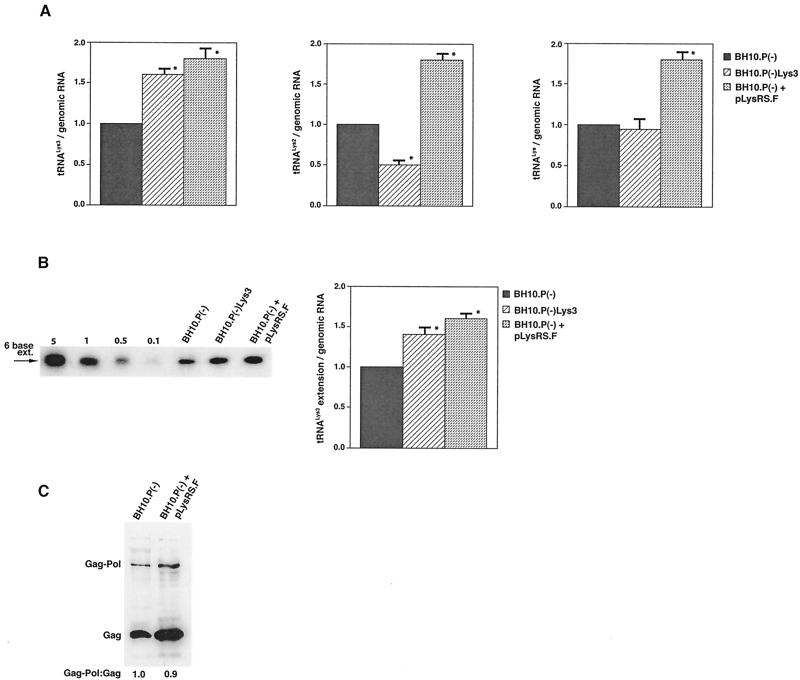
Human immunodeficiency virus type 1 (HIV-1) uses tRNA(Lys)(3) as a primer for reverse transcription and, during viral assembly, this tRNA is selectively packaged into the virus along with the other major tRNA(Lys), tRNA(Lys)(3). Increasing the cytoplasmic concentration of tRNA(Lys)(3) through transfection of cells with a plasmid containing both HIV-1 proviral DNA and a tRNA(Lys)(3) gene results in a greater incorporation of tRNA(Lys)(3) into virions, which is accompanied by increased annealing of tRNA(Lys)(3) to the viral genome and increased infectivity of the viral population. Increased viral tRNA(Lys)(3) is accompanied by decreased viral tRNA(Lys)(3), with the total tRNA(Lys)/virion and the GagPol/Gag ratios remaining unchanged. Viral tRNA(Lys) can be doubled, with increases in both tRNA(Lys)(3) and tRNA(Lys)(1,2) concentrations, by overexpressing lysyl tRNA synthetase. This also results in increased tRNA(Lys)(3) annealing to the viral RNA and increased viral infectivity but, again, no change in the GagPol/Gag ratio was observed. This result indicates that GagPol, whose interaction is required during packaging, is not a limiting factor during tRNA(Lys) incorporation into HIV-1, whereas LysRS is.
Figures
References
-
- Harada, F., G. G. Peters, and J. E. Dahlberg. 1979. The primer tRNA for Moloney murine leukemia virus DNA synthesis: nucleotide sequence and aminoacylation of tRNAPro. J. Biol. Chem. 254:10979-10985. - PubMed
-
- Harada, F., R. C. Sawyer, and J. E. Dahlberg. 1975. A primer RNA for initiation of in vitro Rous sarcoma virus DNA synthesis: nucleotide sequence and amino acid acceptor activity. J. Biol. Chem. 250:3487-3497. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
