

Nucleoside analog resistance caused by insertions in the fingers of human immunodeficiency virus type 1 reverse transcriptase involves ATP-mediated excision
- PMID: 12186898
- PMCID: PMC136461
- DOI: 10.1128/jvi.76.18.9143-9151.2002
Nucleoside analog resistance caused by insertions in the fingers of human immunodeficiency virus type 1 reverse transcriptase involves ATP-mediated excision
Abstract
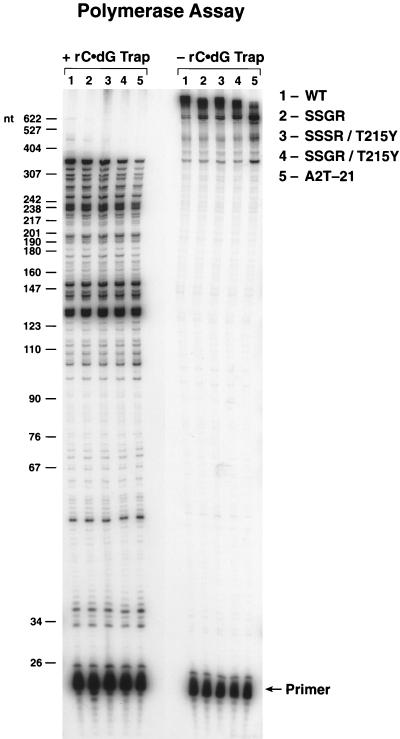
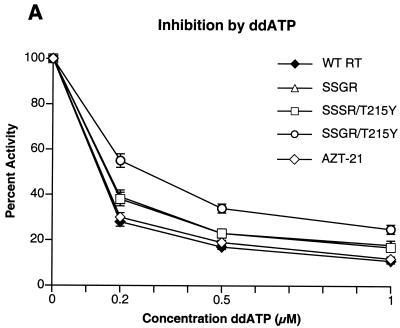
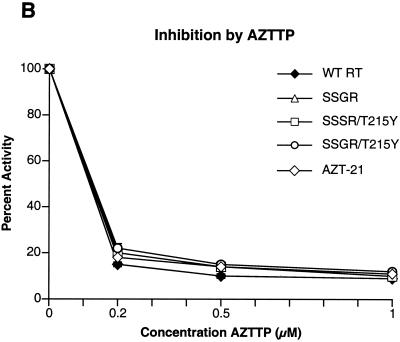
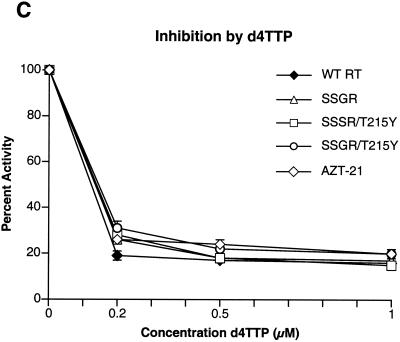
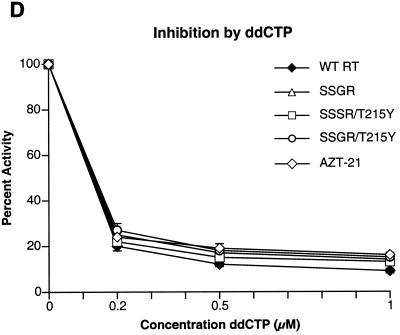
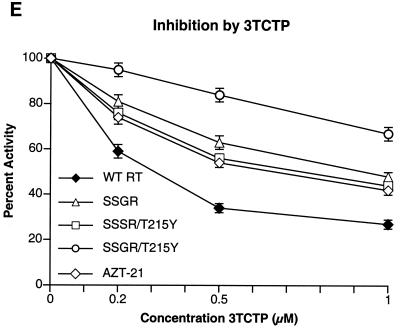
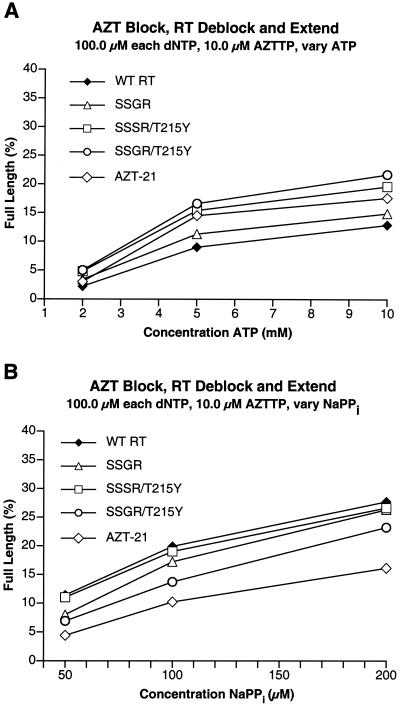
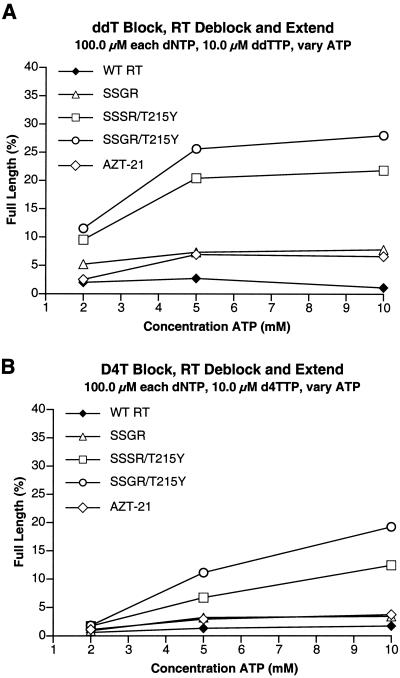
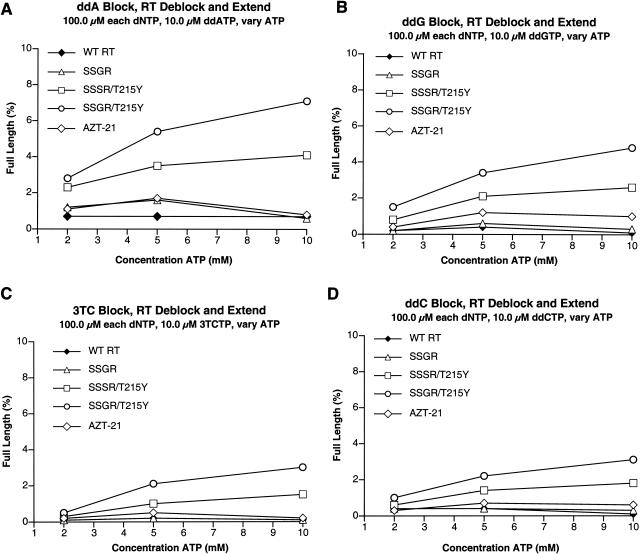
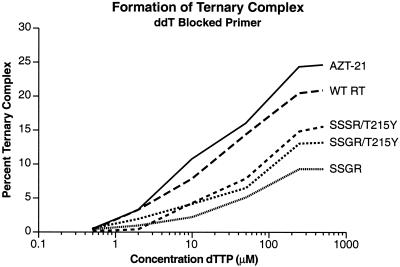
Although anti-human immunodeficiency virus type 1 (HIV-1) therapy has prolonged the lives of patients, drug resistance is a significant problem. Of particular concern are mutations that cause cross-resistance to a particular class of drugs. Among the mutations that cause resistance to several nucleoside analogs are the insertion of amino acids in the fingers subdomain of HIV-1 reverse transcriptase (RT) at positions 69 and 70. These insertions are usually associated with changes in the flanking amino acids and with a change to F or Y at position 215. We have proposed that the T215F/Y mutation makes the binding of ATP to HIV-1 RT more effective, which increases the excision of 3-azido-3'-deoxythymidine-5'-monophosphate (AZTMP) in vitro and increases zidovudine (AZT) resistance in vivo. Although the mechanism of AZT resistance involves enhanced excision, resistance to 3TC involves a block to incorporation of the analog. We measured the effects of fingers insertion mutations on the misincorporation and excision of several nucleoside analogs. RT variants with the amino acid insertions in the fingers and T215Y have a decreased level of misincorporation of ddATP and 3TCTP. These mutants also have the ability to excise AZTMP by ATP-dependent pyrophosphorylysis. However, unlike the classic AZT resistance mutations (M41L/D67N/K70R/T215Y or F/K219E or Q), the combination of the amino acid insertions in the fingers and the T215Y mutation allows efficient excision of ddTMP and d4TMP, even when relatively high levels of deoxynucleoside triphosphates are present in the reaction. Although the dideoxynucleoside analogs of other nucleosides were excised more slowly than AZTMP, ddTMP, and d4TMP, the mutants with the fingers insertion and T215Y excised all of the nucleoside analogs that were tested more efficiently than wild-type RT or a mutant RT carrying the classical AZT resistance mutations. In the ternary complex (RT/template-primer/dNTP), the presence of the bound dNTP prevents the end of the primer from gaining access to the nucleotide binding site (N site) where excision occurs. Gel shift analysis showed that the amino acid insertions in the fingers destabilized the ternary complex compared to wild-type HIV-1 RT. If the ternary complex is unstable, the end of the primer can gain access to the N site and excision can occur. This could explain the enhanced excision of the nucleoside analogs.
Figures
References
-
- Ahluwalia, G., D. A. Cooney, H. Mitsuya, A. Fridland, K. P. Flora, Z. Hao, M. Dalal, S. Broder, and D. G. Johns. 1987. Initial studies on the cellular pharmacology of 2′,3′-dideoxyinosine, an inhibitor of HIV infectivity. Biochem. Pharmacol. 36:3797-3800. - PubMed
-
- Andréoletti, L., L. Weiss, A. Si-Mohamed, C. Piketty, T. Prazuck, G. Calamy, J.-E. Malkin, M. Matta, F.-X. Mbopi-Kéou, F. Clavel, M. D. Kazatchkine, and L. Bélec. 2002. Multidrug-resistant HIV-1 RNA and proviral DNA variants harboring new dipeptide insertions in the reverse transcriptase pol gene. J. Acquir. Immune Defic. Syndr. 29:102-104. - PubMed
-
- Arion, D., N. Kaushik, S. McCormick, G. Borkow, and M. A. Parniak. 1998. Phenotypic mechanism of HIV-1 resistance to 3′-azido-3′-deoxythymidine (AZT): increased polymerization processivity and enhanced sensitivity to pyrophosphate of the mutant viral reverse transcriptase. Biochemistry 37:15908-15917. - PubMed
-
- Bonfanti, P., I. Faggion, S. L. S. Catamancio, M. Violin, C. Balotta, and S. Rusconi. 2000. Response to antiretroviral therapy in a patient with an uncommon codon 69 insertion in the human immunodeficiency virus type 1 reverse transcriptase. Antimicrob. Agents Chemother. 44:1767-1768. - PMC - PubMed
-
- Boyer, P. L., J. Lisziewicz, F. Lori, and S. H. Hughes. 1999. Analysis of amino acid insertion mutations in the fingers subdomain of HIV-1 reverse transcriptase. J. Mol. Biol. 286:995-1008. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
