

Coupling of termination, 3' processing, and mRNA export
- PMID: 12192043
- PMCID: PMC135649
- DOI: 10.1128/MCB.22.18.6441-6457.2002
Coupling of termination, 3' processing, and mRNA export
Abstract
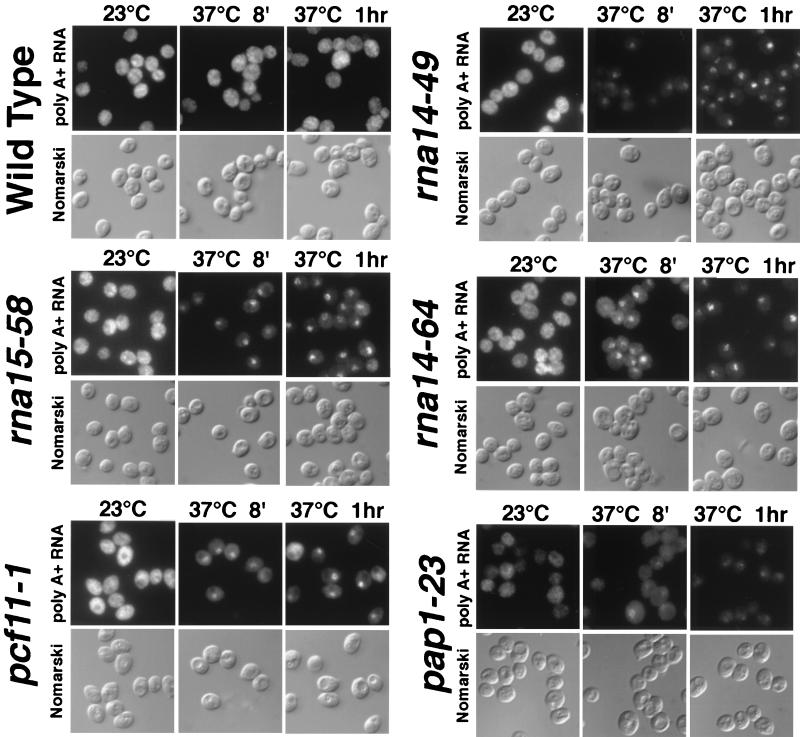
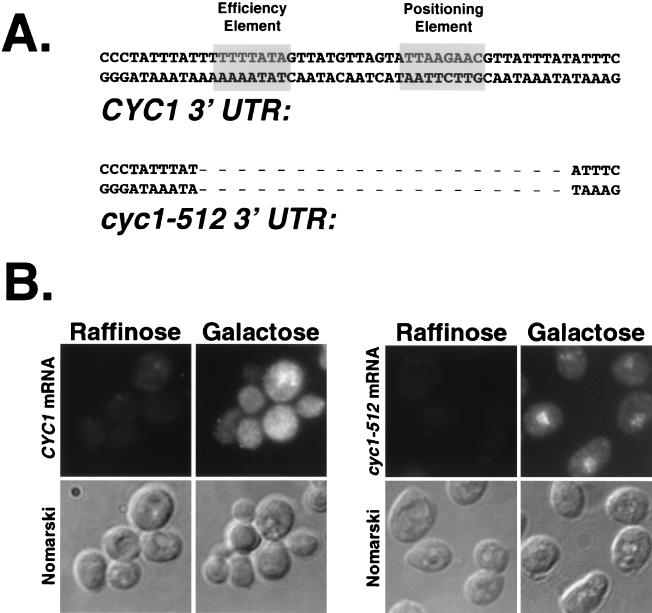
In a screen to identify genes required for mRNA export in Saccharomyces cerevisiae, we isolated an allele of poly(A) polymerase (PAP1) and novel alleles encoding several other 3' processing factors. Many newly isolated and some previously described mutants (rna14-48, rna14-49, rna14-64, rna15-58, and pcf11-1 strains) are defective in polymerase II (Pol II) termination but, interestingly, retain the ability to polyadenylate these improperly processed transcripts at the nonpermissive temperature. Deletion of the cis-acting sequences required to couple 3' processing and termination also produces transcripts that fail to exit the nucleus, suggesting that all of these processes (cleavage, termination, and export) are coupled. We also find that several but not all mRNA export mutants produce improperly 3' processed transcripts at the nonpermissive temperature. 3' maturation defects in mRNA export mutants include improper Pol II termination and/or the previously characterized hyperpolyadenylation of transcripts. Importantly, not all mRNA export mutants have defects in 3' processing. The similarity of the phenotypes of some mRNA export mutants and 3' processing mutants indicates that some factors from each process may mechanistically interact to couple mRNA processing and export. Consistent with this assumption, we present evidence that Xpo1p interacts in vivo with several 3' processing factors and that the addition of recombinant Xpo1p to in vitro processing reaction mixtures stimulates 3' maturation. Of the core 3' processing factors tested (Rna14p, Rna15p, Pcf11p, Hrp1p, Fip1p, and Cft1p), only Hrp1p shuttles. Overexpression of Rat8p/Dbp5p suppresses both 3' processing and mRNA export defects found in xpo1-1 cells.
Figures





References
-
- Afonina, E., R. Stauber, and G. N. Pavlakis. 1998. The human poly(A)-binding protein 1 shuttles between the nucleus and the cytoplasm. J. Biol. Chem. 273:13015-13021. - PubMed
-
- Aitchison, J. D., G. Blobel, and M. P. Rout. 1996. Kap104p: a karyopherin involved in the nuclear transport of messenger RNA binding proteins. Science 274:624-627. - PubMed
-
- Allen, N. P., L. Huang, A. Burlingame, and M. Rexach. 2001. Proteomic analysis of nucleoporin interacting proteins. J. Biol. Chem. 276:29268-29274. - PubMed
-
- Amberg, D. C., A. L. Goldstein, and C. N. Cole. 1992. Isolation and characterization of RAT1: an essential gene of Saccharomyces cerevisiae required for the efficient nucleocytoplasmic trafficking of mRNA. Genes Dev. 6:1173-1189. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases