

Microtubule-dependent subcellular redistribution of the transcriptional coactivator p/CIP
- PMID: 12192059
- PMCID: PMC135647
- DOI: 10.1128/MCB.22.18.6611-6626.2002
Microtubule-dependent subcellular redistribution of the transcriptional coactivator p/CIP
Abstract
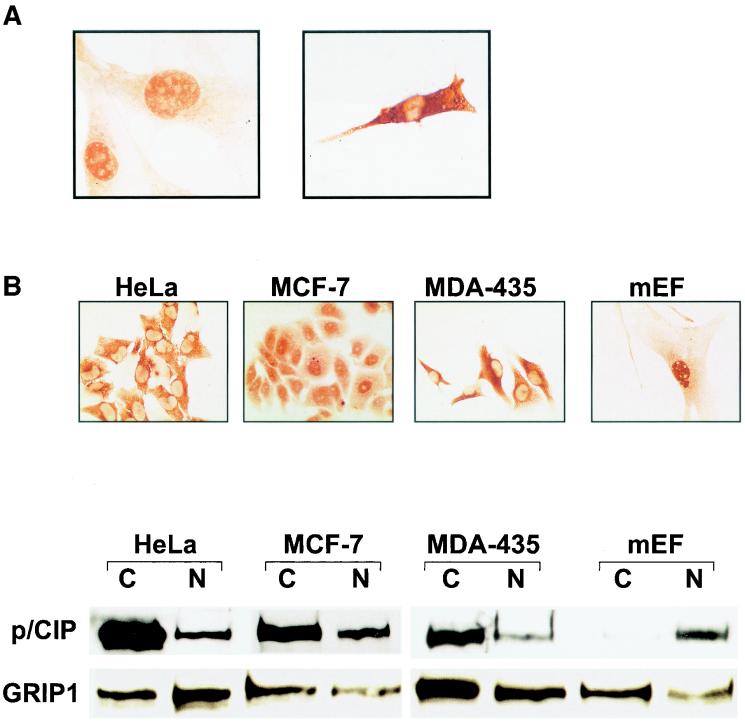
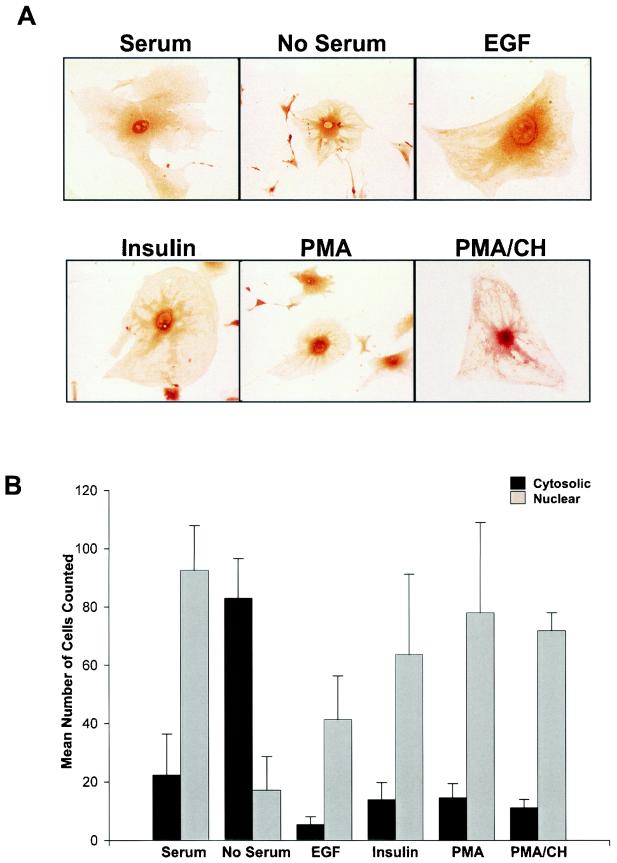
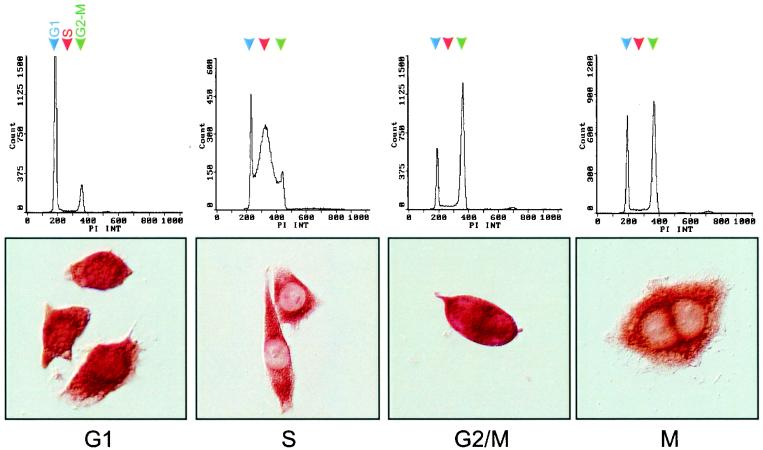
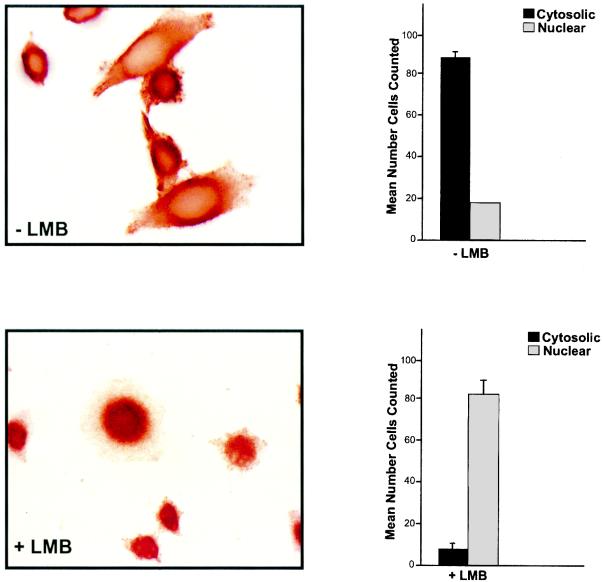
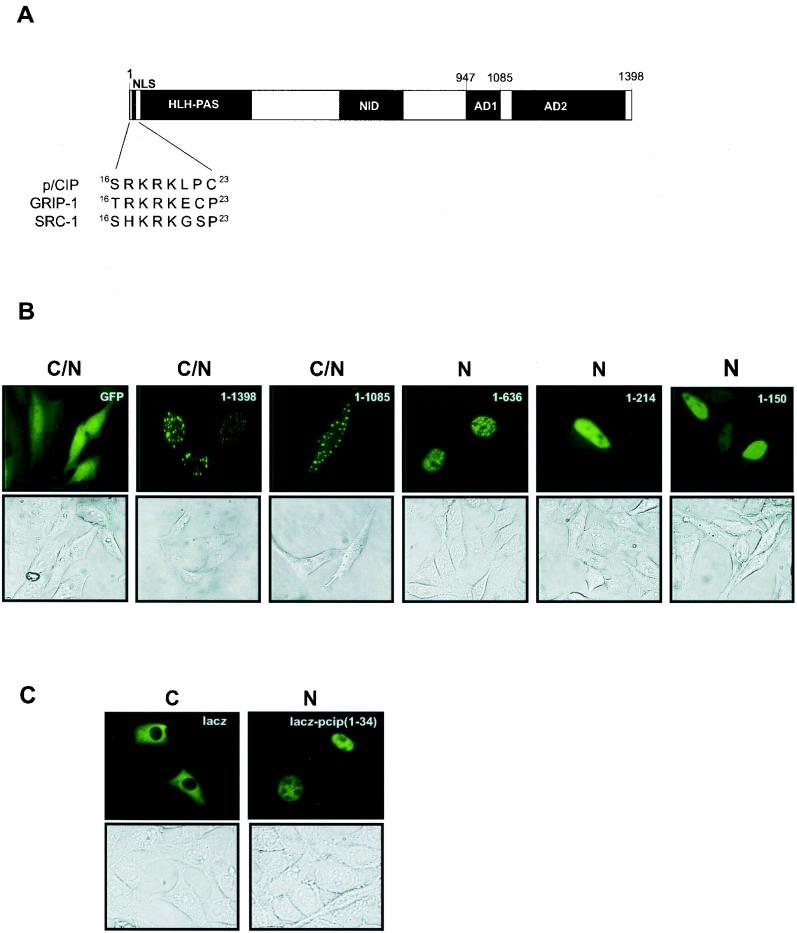
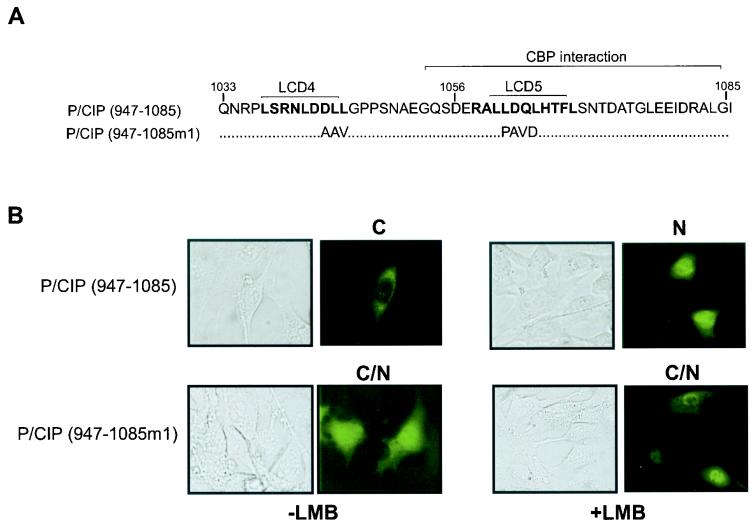
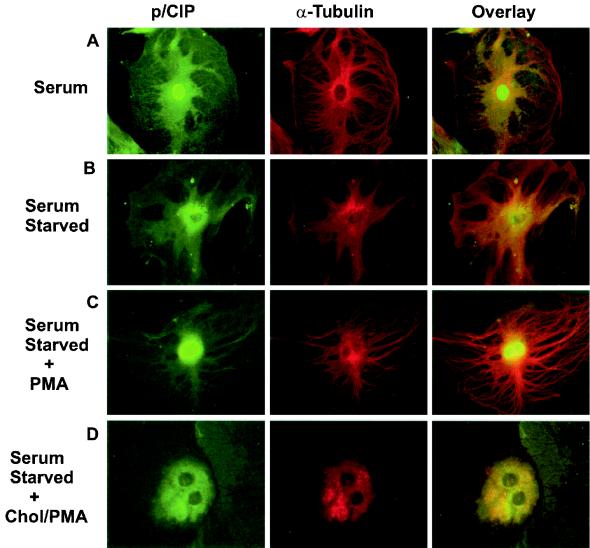
The transcriptional coactivator p/CIP is a member of a family of nuclear receptor coactivator/steroid receptor coactivator (NCoA/SRC) proteins that mediate the transcriptional activities of nuclear hormone receptors. We have found that p/CIP is predominantly cytoplasmic in a large proportion of cells in various tissues of the developing mouse and in a number of established cell lines. In mouse embryonic fibroblasts, serum deprivation results in the redistribution of p/CIP to the cytoplasmic compartment and stimulation with growth factors or tumor-promoting phorbol esters promotes p/CIP shuttling into the nucleus. Cytoplasmic accumulation of p/CIP is also cell cycle dependent, occurring predominantly during the S and late M phases. Leptomycin B (LMB) treatment results in a marked nuclear accumulation, suggesting that p/CIP undergoes dynamic nuclear export as well as import. We have identified a strong nuclear import signal in the N terminus of p/CIP and two leucine-rich motifs in the C terminus that resemble CRM-1-dependent nuclear export sequences. When fused to green fluorescent protein, the nuclear export sequence region is cytoplasmic and is retained in the nucleus in an LMB-dependent manner. Disruption of the leucine-rich motifs prevents cytoplasmic accumulation. Furthermore, we demonstrate that cytoplasmic p/CIP associates with tubulin and that an intact microtubule network is required for intracellular shuttling of p/CIP. Immunoaffinity purification of p/CIP from nuclear and cytosolic extracts revealed that only nuclear p/CIP complexes possess histone acetyltransferase activity. Collectively, these results suggest that cellular compartmentalization of NCoA/SRC proteins could potentially regulate nuclear hormone receptor-mediated events as well as integrating signals in response to different environmental cues.
Figures
Similar articles
-
The coactivator p/CIP/SRC-3 facilitates retinoic acid receptor signaling via recruitment of GCN5.J Biol Chem. 2003 Oct 10;278(41):39402-12. doi: 10.1074/jbc.M307832200. Epub 2003 Jul 28. J Biol Chem. 2003. PMID: 12885766
-
The activity and stability of the transcriptional coactivator p/CIP/SRC-3 are regulated by CARM1-dependent methylation.Mol Cell Biol. 2007 Jan;27(1):120-34. doi: 10.1128/MCB.00815-06. Epub 2006 Oct 16. Mol Cell Biol. 2007. PMID: 17043108 Free PMC article.
-
The transcriptional co-activator p/CIP binds CBP and mediates nuclear-receptor function.Nature. 1997 Jun 12;387(6634):677-84. doi: 10.1038/42652. Nature. 1997. PMID: 9192892
-
Review of the in vivo functions of the p160 steroid receptor coactivator family.Mol Endocrinol. 2003 Sep;17(9):1681-92. doi: 10.1210/me.2003-0116. Epub 2003 Jun 12. Mol Endocrinol. 2003. PMID: 12805412 Review.
-
The SRC family of nuclear receptor coactivators.Gene. 2000 Mar 7;245(1):1-11. doi: 10.1016/s0378-1119(00)00024-x. Gene. 2000. PMID: 10713439 Review.
Cited by
-
Overexpression of AIB1 correlates inversely with E-cadherin expression in pancreatic adenocarcinoma and may promote lymph node metastasis.Int J Clin Oncol. 2014 Apr;19(2):319-24. doi: 10.1007/s10147-013-0549-2. Epub 2013 Mar 30. Int J Clin Oncol. 2014. PMID: 23542947
-
Intrinsically Disordered SRC-3/AIB1 Protein Undergoes Homeostatic Nuclear Extrusion by Nuclear Budding While Ectopic Expression Induces Nucleophagy.Cells. 2019 Oct 19;8(10):1278. doi: 10.3390/cells8101278. Cells. 2019. PMID: 31635050 Free PMC article.
-
Redundant enhancement of mouse constitutive androstane receptor transactivation by p160 coactivator family members.Arch Biochem Biophys. 2007 Dec 1;468(1):49-57. doi: 10.1016/j.abb.2007.09.005. Epub 2007 Sep 18. Arch Biochem Biophys. 2007. PMID: 17950690 Free PMC article.
-
The nuclear coactivator amplified in breast cancer 1 maintains tumor-initiating cells during development of ductal carcinoma in situ.Oncogene. 2014 Jun 5;33(23):3033-42. doi: 10.1038/onc.2013.263. Epub 2013 Jul 15. Oncogene. 2014. PMID: 23851504 Free PMC article.
-
Regulation of the BRCA1 gene by an SRC3/53BP1 complex.BMC Biochem. 2011 Sep 13;12:50. doi: 10.1186/1471-2091-12-50. BMC Biochem. 2011. PMID: 21914189 Free PMC article.
References
-
- Anafi, M., Y. F. Yang, N. A. Barlev, M. V. Govindan, S. L. Berger, T. R. Butt, and P. G. Walfish. 2000. GCN5 and ADA adaptor proteins regulate triiodothyronine/GRIP1 and SRC-1 coactivator-dependent gene activation by the human thyroid hormone receptor. Mol. Endocrinol. 14:718-732. - PubMed
-
- Anzick, S. L., J. Kononen, R. L. Walker, D. O. Azorsa, M. M. Tanner, X. Y. Guan, G. Sauter, O. P. Kallioniemi, J. M. Trent, and P. S. Meltzer. 1997. AIB1, a steroid receptor coactivator amplified in breast and ovarian cancer. Science 277:965-968. - PubMed
-
- Braiman, L., L. Sheffi-Friedman, A. Bak, T. Tennenbaum, and S. R. Sampson. 1999. Tyrosine phosphorylation of specific protein kinase C isoenzymes participates in insulin stimulation of glucose transport in primary cultures of rat skeletal muscle. Diabetes 48:1922-1929. - PubMed
-
- Chen, D., H. Ma, H. Hong, S. S. Koh, S. M. Huang, B. T. Schurter, D. W. Aswad, and M. R. Stallcup. 1999. Regulation of transcription by a protein methyltransferase. Science 284:2174-2177. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous