

Surface adhesion of fusion proteins containing the hydrophobins HFBI and HFBII from Trichoderma reesei
- PMID: 12192081
- PMCID: PMC2373586
- DOI: 10.1110/ps.0207902
Surface adhesion of fusion proteins containing the hydrophobins HFBI and HFBII from Trichoderma reesei
Abstract
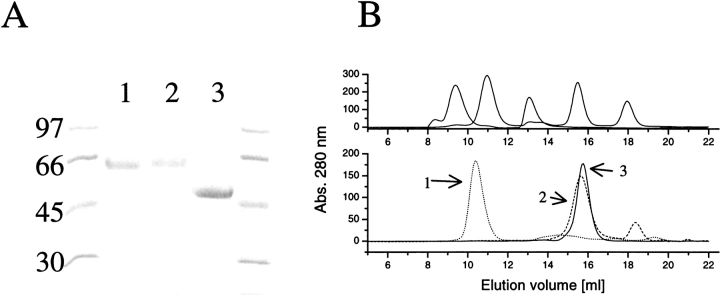
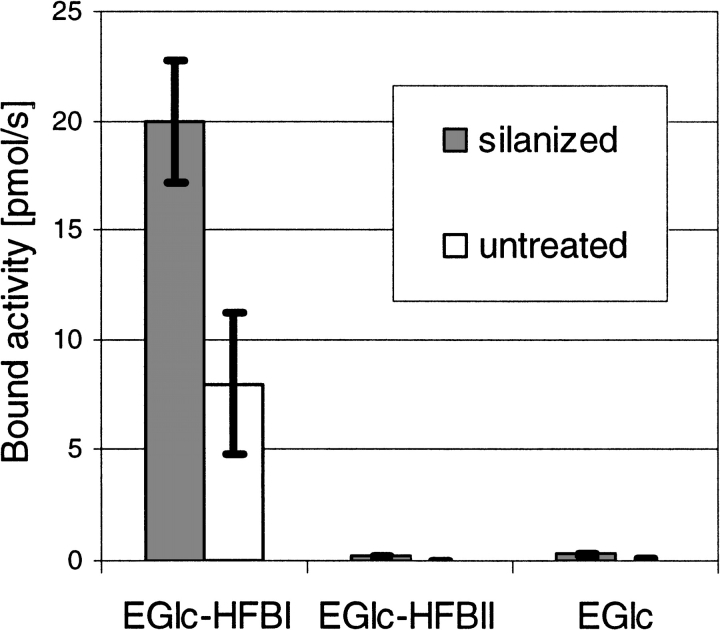
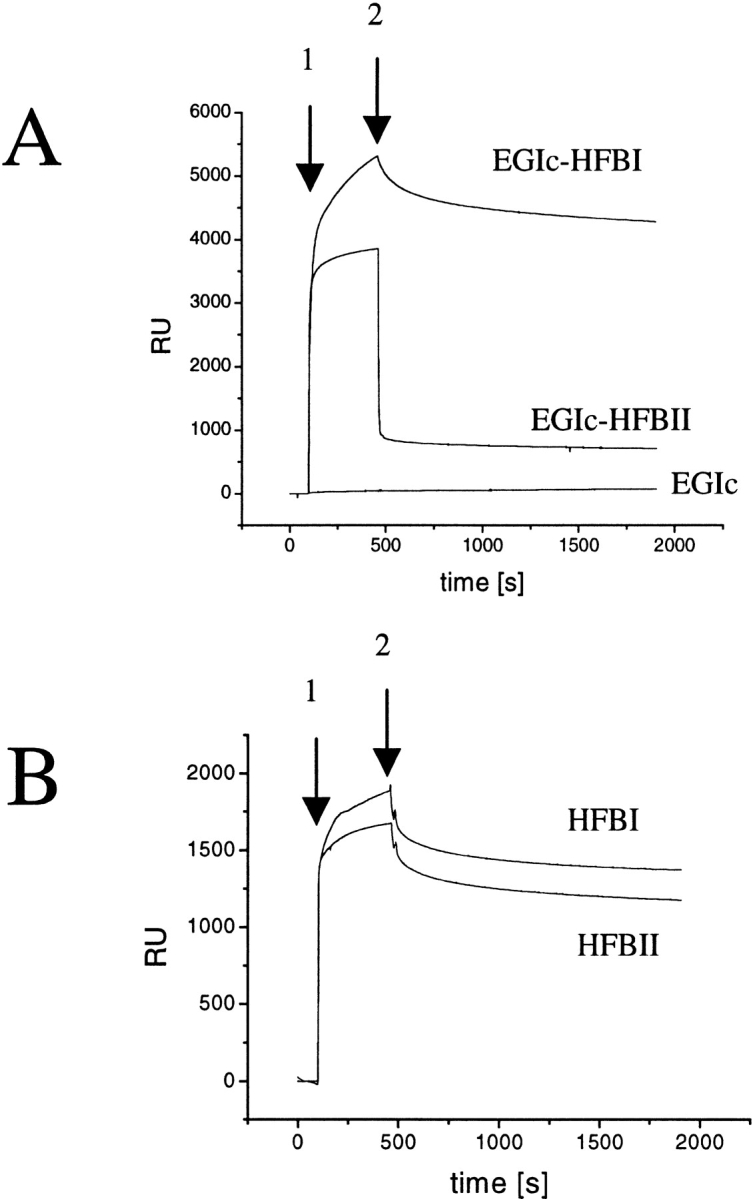
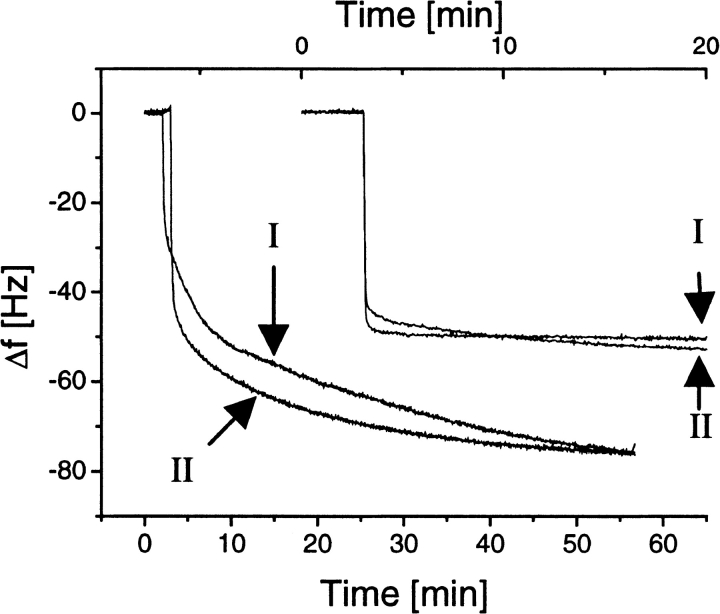
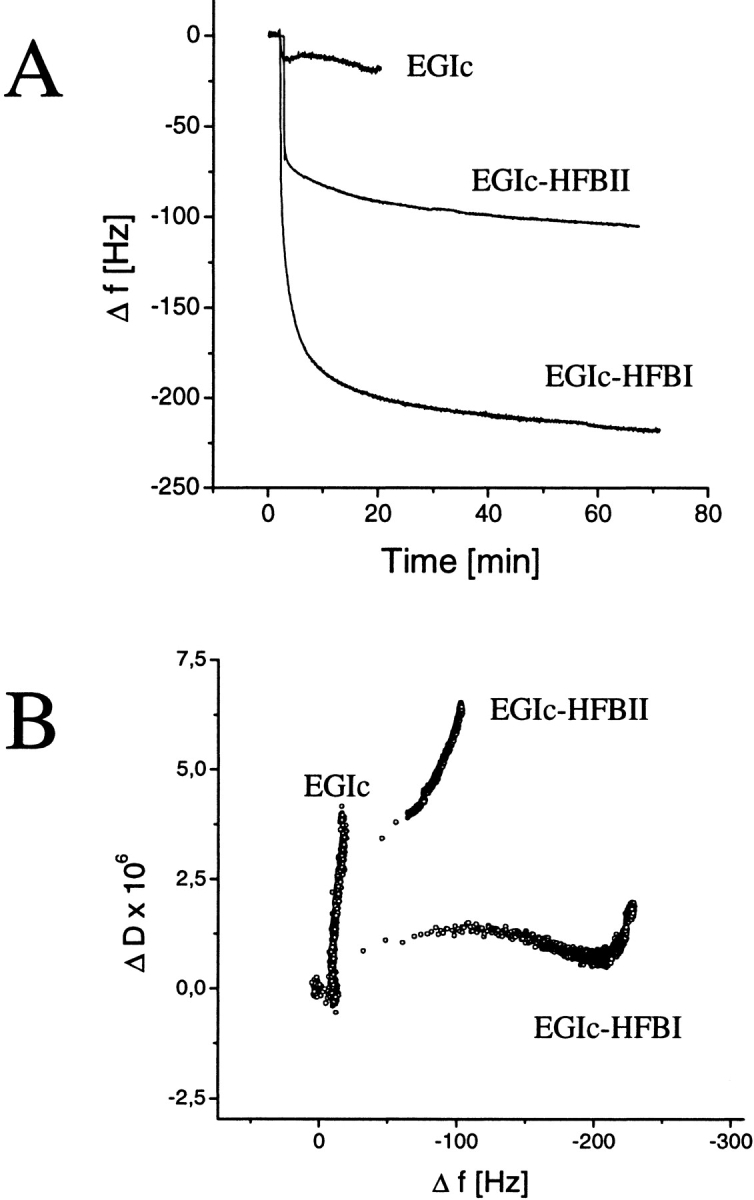
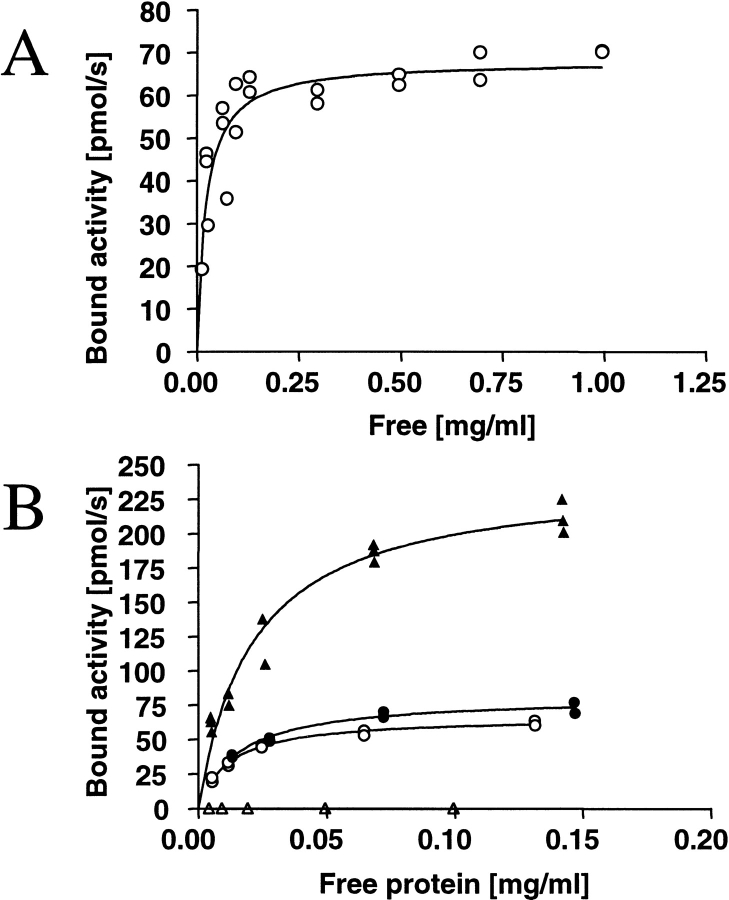
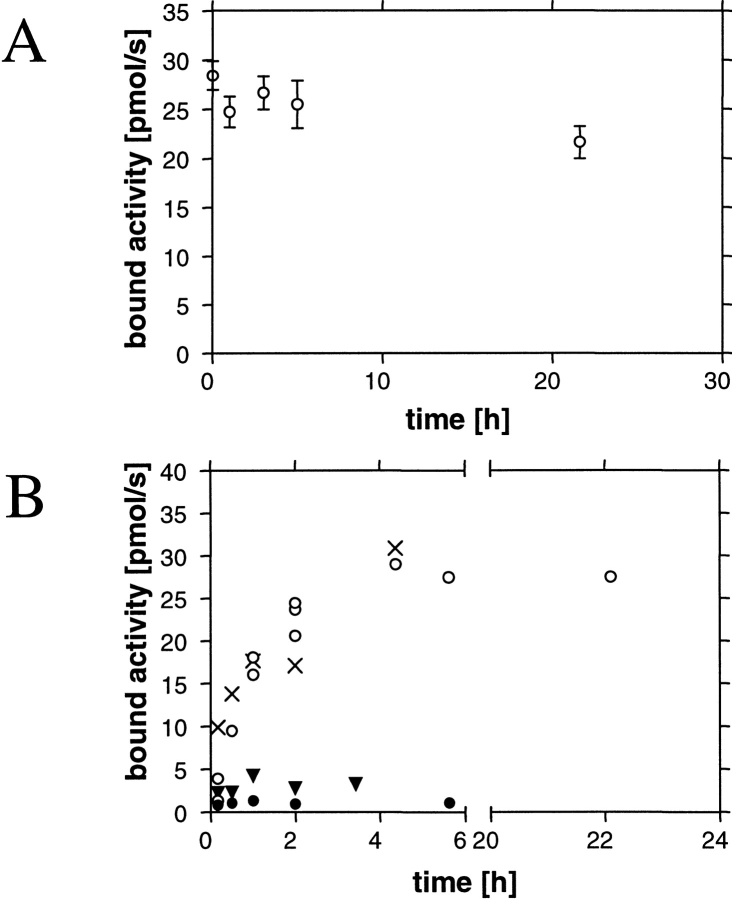
Hydrophobins are surface-active proteins produced by filamentous fungi, where they seem to be ubiquitous. They have a variety of roles in fungal physiology related to surface phenomena, such as adhesion, formation of surface layers, and lowering of surface tension. Hydrophobins can be divided into two classes based on the hydropathy profile of their primary sequence. We have studied the adhesion behavior of two Trichoderma reesei class II hydrophobins, HFBI and HFBII, as isolated proteins and as fusion proteins. Both hydrophobins were produced as C-terminal fusions to the core of the hydrolytic enzyme endoglucanase I from the same organism. It was shown that as a fusion partner, HFBI causes the fusion protein to efficiently immobilize to hydrophobic surfaces, such as silanized glass and Teflon. The properties of the surface-bound protein were analyzed by the enzymatic activity of the endoglucanase domain, by surface plasmon resonance (Biacore), and by a quartz crystal microbalance. We found that the HFBI fusion forms a tightly bound, rigid surface layer on a hydrophobic support. The HFBI domain also causes the fusion protein to polymerize in solution, possibly to a decamer. Although isolated HFBII binds efficiently to surfaces, it does not cause immobilization as a fusion partner, nor does it cause polymerization of the fusion protein in solution. The findings give new information on how hydrophobins function and how they can be used to immobilize fusion proteins.
Figures
Similar articles
-
Interaction and comparison of a class I hydrophobin from Schizophyllum commune and class II hydrophobins from Trichoderma reesei.Biomacromolecules. 2006 Apr;7(4):1295-301. doi: 10.1021/bm050676s. Biomacromolecules. 2006. PMID: 16602752
-
Behavior of Trichoderma reesei hydrophobins in solution: interactions, dynamics, and multimer formation.Biochemistry. 2006 Jul 18;45(28):8590-8. doi: 10.1021/bi060620y. Biochemistry. 2006. PMID: 16834333
-
The hydrophobins HFBI and HFBII from Trichoderma reesei showing efficient interactions with nonionic surfactants in aqueous two-phase systems.Biomacromolecules. 2001 Summer;2(2):511-7. doi: 10.1021/bm0001493. Biomacromolecules. 2001. PMID: 11749214
-
Hydrophobins: the protein-amphiphiles of filamentous fungi.FEMS Microbiol Rev. 2005 Nov;29(5):877-96. doi: 10.1016/j.femsre.2005.01.004. Epub 2005 Feb 21. FEMS Microbiol Rev. 2005. PMID: 16219510 Review.
-
Applications of hydrophobins: current state and perspectives.Appl Microbiol Biotechnol. 2015 Feb;99(4):1587-97. doi: 10.1007/s00253-014-6319-x. Epub 2015 Jan 8. Appl Microbiol Biotechnol. 2015. PMID: 25564034 Review.
Cited by
-
Aggregation and self-assembly of hydrophobins from Trichoderma reesei: low-resolution structural models.Biophys J. 2002 Oct;83(4):2240-7. doi: 10.1016/S0006-3495(02)73984-2. Biophys J. 2002. PMID: 12324441 Free PMC article.
-
Fusion to Hydrophobin HFBI Improves the Catalytic Performance of a Cytochrome P450 System.Front Bioeng Biotechnol. 2016 Jul 4;4:57. doi: 10.3389/fbioe.2016.00057. eCollection 2016. Front Bioeng Biotechnol. 2016. PMID: 27458582 Free PMC article.
-
Recent Advances in Fungal Hydrophobin Towards Using in Industry.Protein J. 2015 Aug;34(4):243-55. doi: 10.1007/s10930-015-9621-2. Protein J. 2015. PMID: 26208665 Review.
-
Two novel class II hydrophobins from Trichoderma spp. stimulate enzymatic hydrolysis of poly(ethylene terephthalate) when expressed as fusion proteins.Appl Environ Microbiol. 2013 Jul;79(14):4230-8. doi: 10.1128/AEM.01132-13. Epub 2013 May 3. Appl Environ Microbiol. 2013. PMID: 23645195 Free PMC article.
-
A novel hydrophobin encoded by hgfII from Grifola frondosa exhibiting excellent self-assembly ability.Front Microbiol. 2022 Sep 9;13:990231. doi: 10.3389/fmicb.2022.990231. eCollection 2022. Front Microbiol. 2022. PMID: 36160239 Free PMC article.
References
-
- Alfthan, K. 1998. Surface plasmon resonance biosensors as a tool in antibody engineering. Biosens. Bioelectron. 13 653–663. - PubMed
-
- Aro, N., Saloheimo, A., Ilmen, M., and Penttila, M. 2001. ACEII, a novel transcriptional activator involved in regulation of cellulase and xylanase genes of Trichoderma reesei. J. Biol. Chem. 276 24309–24314. - PubMed
-
- Carpenter, C.E., Mueller, R J., Kazmierczak, P., Zhang, L., Villalon, D.K., and Van Alfen, N.K. 1992. Effect of a virus on accumulation of a tissue-specific cell-surface protein of the fungus Cryphonectria (Endothia) parasitica. Mol. Plant Microbe Interact. 5 55–61. - PubMed
-
- Collen, A., Persson, J., Linder, M., Nakari-Setälä, T., Penttilä, M., Tjerneld, F., and Sivars, U. 2002. A novel two-step extraction method with detergent/polymer systems for primary recovery of the fusion protein endoglucanase I-hydrophobin I. Biochim. Biophys. Acta. 1569 139–150. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
